



Description of a new species of Protodinychus (Mesostigmata: Protodinychidae), and a key to deutonymphs of the genus
Kazemi, Shahrooz  1
and Klompen, Hans
2
1
and Klompen, Hans
2
1✉ Department of Biodiversity, Institute of Science and High Technology and Environmental Sciences, Graduate University of Advanced Technology, Kerman, Iran.
2Acarology Collection, Department of Evolution, Ecology, and Organismal Biology, Ohio State University, Columbus, Ohio, United States of America.
2022 - Volume: 62 Issue: 3 pages: 585-596
https://doi.org/10.24349/6gmw-wgodZooBank LSID: 1DDF09BC-4903-4331-88E9-88E4306B0BB1
Original research
Keywords
Abstract
Introduction
The mesostigmatid lineage Uropodina or turtle mites forms an easy recognizable, diverse and globally distributed grouping. Even so, many aspects of the systematics and biology of the group are still unclear. For example, the systematic position of the entire Uropodina is not settled, with views ranging from association with Microgyniina and Sejina (Johnston in Norton et al. 1993) to a sistergroup relationship with the combined lineage of Sejina plus Gamasina (Klompen 2000; Klompen et al. 2007). The Thinozerconoidea, families Thinozerconidae (1 described species) and Protodinychidae (3 described species), are key taxa in this discussion. Evans (1957) and Hirschmann (1979b) did not consider Thinozercon Evans, 1957 a uropodid, and Wisniewski and Hirschmann (1993) never even mention Thinozercon or Protodinychus Evans, 1957 in their catalog of Uropodina, but most researchers list them as probable sistergroup to all other Uropodina (Johnston 1961; Athias-Binche and Evans 1981; Karg 1989; Lindquist et al. 2009). Improved understanding of Thinozerconoidea may therefore be critical to understanding of higher order relationships in Mesostigmata, as well as understanding of morphological and biological evolution within Uropodina.
One of several oddities regarding Thinozerconoidea concerns their geographic distribution. Thinozercon was described from the coast of Ireland, with additional reports from the coast of Wales (Pugh et al. 1987) and from Oregon (listed as an unidentified thionozerconid in Krantz and Ainscough 1990). Within Protodinychidae, Protodinychus punctatus Evans, 1957 was described from England, and has been reported from Scotland, Ireland (Athias-Binche and Evans 1981), The Netherlands (Siepel et al. 2018), Hungary (Kontschán 2015), Poland (Napierala and Bloszyk 2013), and Latvia (Salmane and Spungis 2015). Protodinychus evansi Huțu & Călugăr, 2002 and P. ainscoughi Huțu & Călugăr, 2002 were described based on specimens collected in Canada (Johnston 1961; Huțu and Călugăr 2002) and Romania (Huțu and Călugăr 2002; Kontschán 2014). Chaudhury et al. (2005) reported an unknown species of Protodinychus from India, which would be a huge range extension, but the identification of the genus could not be confirmed. The current confirmed distribution is therefore decidedly north temperate, while the main diversity of Uropodina can be described as tropical (main diversity) or cosmopolitan (broad occurrence).
Two recent studies (Babaeian 2016; Hajizadeh et al. 2021), each based on a single specimen, reported the presence of Protodinychus punctatus in Iran, which would represent a significant range extension for the genus and the species. We obtained a third deutonymph from Mazandaran province, Iran. This deutonymph and the Babaeian specimen (the Hajizadeh et al. specimen is conspecific but could not be studied in detail) exhibit characters inconsistent with the description of P. punctatus or any of the other previously described species. Descriptions of new species based on 2–3 immature specimens are generally to be avoided, but seem justified in this case because of the systematic importance of the lineage to which these specimens belong. Second, while previous descriptions of Protodinychus species were adequate for their time, they leave room for improvement, especially in terms of distribution of lyrifissures and glands. The goal of this study is therefore to describe these deutonymphs as a new species, to present additional information on P. ainscoughi, and to provide a key to the deutonymphs of the genus.
Material and methods
All specimens studied were extracted using Berlese-Tullgren funnels and mounted in Hoyer's medium on microscope slides.
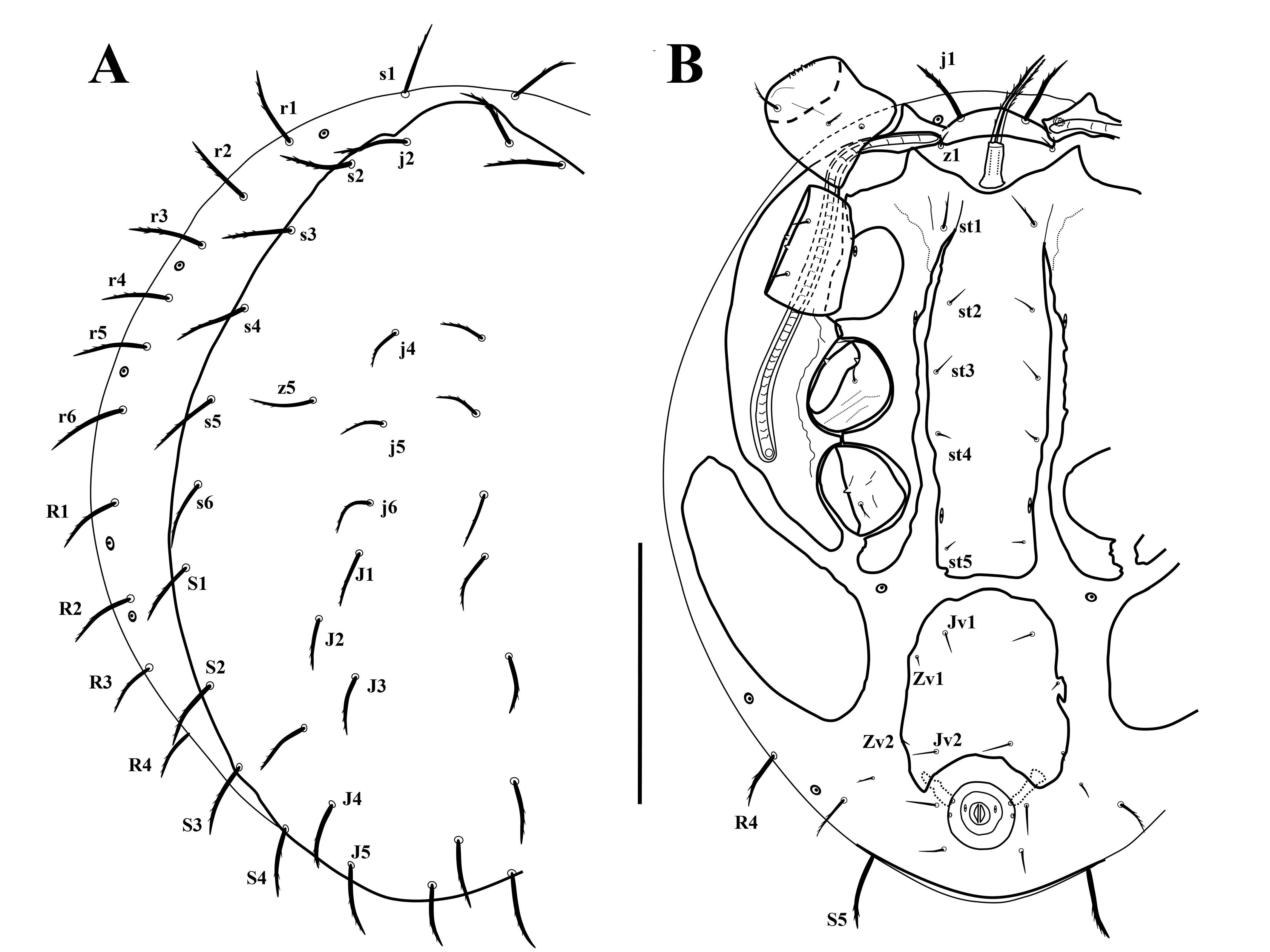
Morphological observations, measurements, and illustrations were made using a Zeiss Axioskop compound microscope with phase contrast optics and a drawing tube. Additional images were prepared using the automated Z-stacking feature of the Nikon NIS Elements package on a Nikon Eclipse 90i (Melville, NY) compound microscope with a PC controlled Ds-5M-U1 digital camera at the Acarology Laboratory of the Ohio State University (OSAL). Measurements were made in micrometers (μm). Dorsal shield length and width were taken from the anterior to the posterior shield margins along the midline and the lateral margins at the broadest level, respectively. The length and width of the sternal and ventral shields were measured from the anterior to posterior margins of the shields along the midline and from the lateral margins of the shields at the broadest point, respectively. The length of the second cheliceral segment was measured from the base of the segment to the apex of the fixed digit, the length of the movable digit from the base to the apex of the chela. Leg lengths were taken from the base of the coxa to the apex of the tarsus, excluding the pretarsus. Notation for setae on the dorsal and ventral idiosoma follows Athias-Binche and Evans (1981), and palp and leg chaetotaxy follow Evans (1963, 1972).
Family Protodinychidae Evans, 1957
Genus Protodinychus Evans, 1957
Type species: Protodinychus punctatus Evans, 1957: 239
Protodinychus ahangarani n. sp.
ZOOBANK: 737EAAD2-77D1-4675-AE70-C9AF1F9CD077 ![]()
(Figures 1–4E)


Diagnosis (deutonymph)
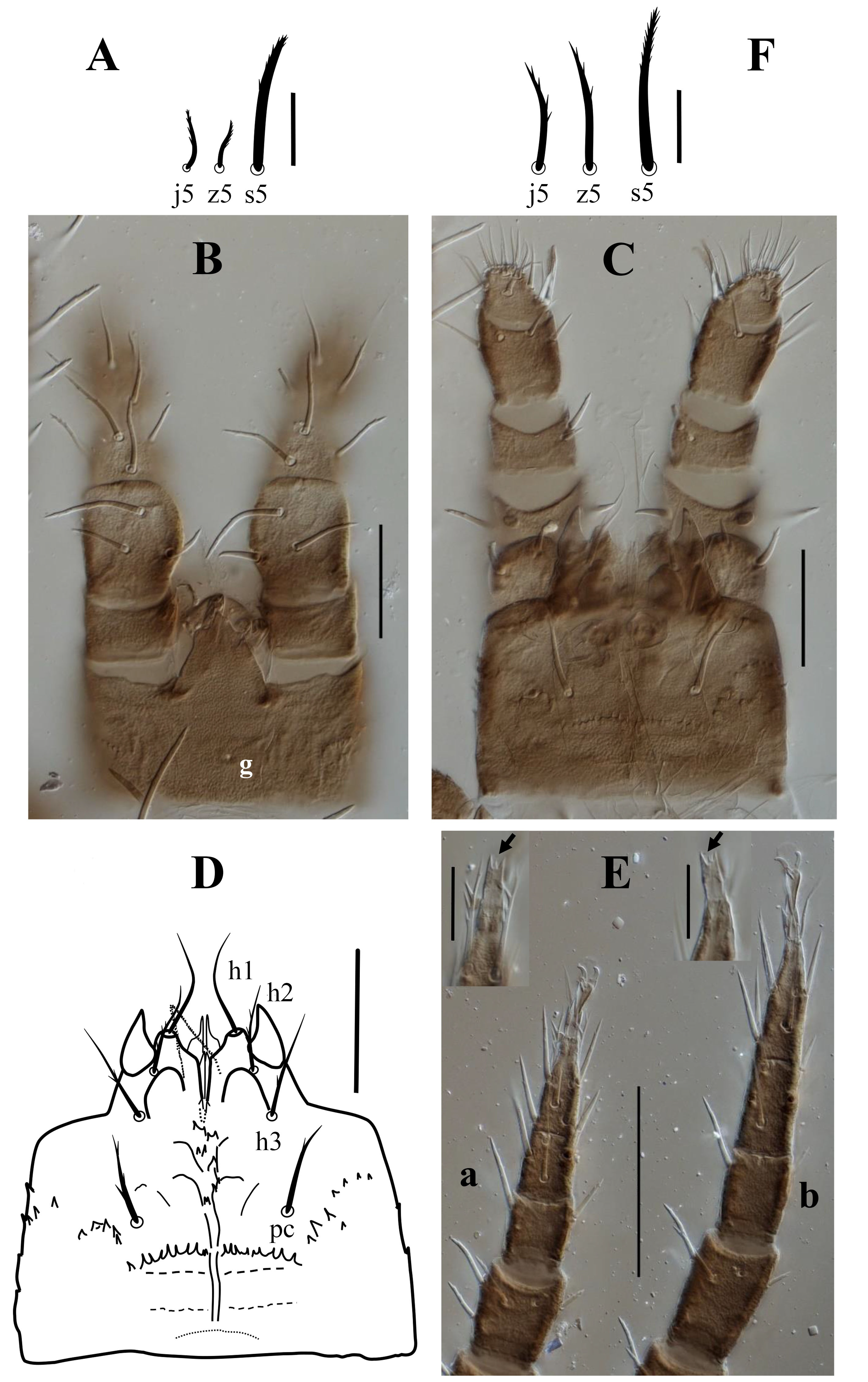
Dorsal shield with 20 pairs of setae; setae z5 shorter than s5, and longer than adjacent j-series setae. Dorsolateral soft cuticle with 11 pairs of subequal setae including s1, r1–r6, R1–R4. Setae j1 inserted anteroventrally on well-developed small vertex platelet. Setae j1 and s1 subequal in length. Setae st1 markedly longer than st2–st5. Ventral shield with four pairs of setae; soft cuticle between metapodal and ventrianal shields devoid of setae. Hypostomal setae h2–h3 and pc barbed.
Deutonymph (N=2)
Dorsal idiosoma (Figures 1A, 2, 4A). Idiosoma suboval, 630 long, 500 wide; dorsal shield 587–609 long, 389–397 wide; idiosoma mostly covered with a suboval holonotal shield bearing 20 pairs of setae: j2 58 long, next anterior two pairs of dorsal setae 34–36 long, and posterior dorsal setae 44–54 long; lateral setae 52–62 long; submarginal setae z5 48–50 long (Figure 4A). Shield surface mostly covered with alveolate ornamentation (Figure 2). One pair of setae (j1) inserted on narrow well-developed vertex plate, located ventrally, flanked by one pair of short, smooth setae z1 (15) and one pair of enlarged poroids, surrounded by a narrow sclerotized band. Lateral soft cuticle bearing 11 pairs of subequal setae, including s1 (51–53), and 10 pairs of r-R series setae (52–62); with eight pairs of enlarged poroids surrounded by a narrow sclerotized band. Count includes poroids near vertex plate, five pairs of dorsal poroids, and two posterior ones inserted ventro-lateral. There are additional glands and poroids on the shields on the idiosomal dorsum and venter, but the structure of the shields did not allow us to confirm their strict number. All dorsal setae (except z1) moderately thickened and barbed.
Ventral idiosoma (Figures 1B, 3). Tritosternal base columnar, 31 long, 18 wide at base, 10 wide at apex; with a pair of finely pilose laciniae, 71–76 long. Sternal shield 284–296 long, 95–97 wide at broadest level, shield with subtriangular anterior and truncate posterior margins, anteromedian margin with a U-shaped depression behind base of tritosternum; endopodal strips free from sternal shield over most of their length except for area between coxae I–II; shield bearing five pairs of setae, st1 27–28, longer than st2 18, st3 18–19, st4 13–14, st5 9–11. Exopodal and peritrematal shields fused, large, extending posteriorly behind coxae IV, with an anterior triangular projection anteriad apex of peritreme. Peritreme almost straight, with finely serrate margin; stigma at anterior level of coxa IV; post-stigmatal extension absent. Ventral shield 139 long, and 138–144 wide at broadest level, sub-rectangular, anterior and posterior margins irregularly convex and concave, respectively; shield bearing four pairs of short and smooth setae, anterior (Jv1) (18) and posterior (Jv2) (22–23) median pairs longer than lateral pairs (Zv1, Zv2) (9–11). Anus surrounded by a narrow sclerotized ridge, bearing two pairs of gland pores, anal valves each bearing one poroid. Anterior para-anal setae (25–26) longer than posterior pair (20), both inserted in soft cuticle. Postanal seta absent. With a pair of large metapodal shields. All ventral shields ornamented with small sub-circular protrusions (Figure 3). Ventral soft cuticle bearing one pair of poroids near coxae II and one pair of poroids behind coxae IV, all of them surrounded by a narrow sclerotized band; with two pairs of ventral setae, anterior pair short (11), smooth, located at posterior level of ventral shield, posterior pair longer (17), barbed, at level of anterior para-anal setae (Figures 1B, 3).
Gnathosoma (Figures 4B–4D). Gnathotectum with a subtriangular anterior margin, bearing one pore dorsomedially (Figure 2, 4B). Internal malae short and smooth; corniculi short, horn-like, and simple; hypostomal setae h1 42–47, smooth, h2 33–34, h3 38–40, and pc 36–38, barbed; deutosternal groove with irregular denticles, flanked by lateral transverse denticulate rows posterior to setae pc (Figures 4C–D). Chelicera similar to those of P. ainscoughi (see Figure 5A), second segment of chelicera length 113; fixed digit 32, edentate, with a sensory organ at the tip; movable digit 36, with a short oblique tooth. Palp length 149–152; chaetotaxy 2-4-5-14-15, palptarsus apotele two-tined (Figure 4C).
Legs (Figure 4E). Length of legs I–IV (left and right legs): 556–559, 436–440, 433–442, and 547–549, respectively. All legs with pretarsus and claws; tarsi II–IV each with one small apicoventral plate, bearing 3–4 teeth at apex (Figure 4E, insets). Leg setae mostly moderately long, and lightly pilose; leg chaetotaxy normal for the genus (Evans 1972), setation of coxae I–IV 2, 2, 2, 1, trochanters I–IV 5, 5, 5, 5, femora I–IV 12 (2 3/1, 2/2, 2), 11 (2, 3/1, 2/2, 1), 6 (1, 2/0, 2/1, 0), 6 (1, 2/0, 2/0, 1), genua I–IV 13 (2, 3/1, 2/1, 2), 11 (2, 3/1, 2/1, 2), 10 (2, 3/1, 2/0, 2), 10 (2, 3/1, 3/0, 1), and tibiae I–IV 11 (2, 2/1, 2/2, 2), 9 (2, 1/1, 2/1, 2), 8 (1, 1/1, 2/1, 2), 9 (1, 1/1, 2/1, 1), respectively.
Material examined
Deutonymph #1, collected from soil-litter mixed with cow manure under Prunus cerasifera (Rosaceae) in Dimarun Village, Tonekabon County, Mazandaran Province, Iran (36.6991° N, 50.6586° E), 1920 m asl, 02 June 2010, coll. Y. Ahangaran (specimen in hand). Deutonymph # 2 collected from soil-litter in the Kheyroud-Kenar Forest, Mazandaran Province (36.6172° N, 51.6092° E), coll. E. Babaeian (studied based on published illustrations and photographs). A third deutonymph was collected from ''soil under broad leaves trees'' in Anzali City, Gilan Province (37.1859° N, 49.6634° E) (Hajizadeh et al. 2021).
All specimens were recovered in the same part of Iran (Gilan Province borders Mazandaran Province), specifically the north slope of the Alborz Mountain range, stretching east-west along the southern margin of the Caspian Sea. The north slopes are usually humid and comprise the Hyrcanian forests on the lowest levels, beech forests in the middle, and oak forests in higher areas.
Type depository
Holotype deutonymph (specimen #1), will be deposited in the Acarological Collection, Institute of Science and High Technology and Environmental Sciences, Graduate University of Advanced Technology, Kerman, Iran. Paratype deutonymph (specimen # 2) deposited in the Acarological Collection of the Jalal Afshar Zoological Museum, Department of Plant Protection, Faculty of Agriculture, University of Tehran, Karaj, Iran.
Etymology
The species is named in honor of Yazdanfar Ahangaran (Forests, Rangelands and Watershed Organization, Department of Natural Protection and Conservation, North of Iran, Chalus), for his help in collecting the mite specimen.
Note
Before this paper, Protodinychus comprised three described species. Protodinychus ahangarani n. sp. differs from the other deutonymphs of Protodinychus species by three unique characters: (1) setae s1 long, as long as j1, vs. s1 short, almost as long as z1 in other species; (2) dorsolateral soft cuticle setae long, subequal in length, vs. variable length in other species; and (3) setae st1 markedly longer than other sternal setae, vs. st1 as long as the other sternal setae in the other species of Protodinychus. With four pairs of setae inserted on the ventral shield the new species resembles P. ainscoughi (Figure 6B), but in addition to the unique features mentioned above, it differs in two more characters: (1) setae s1 in P. ainscoughi inserted on the dorsal shield (Figure 6A), vs. off the shield in the new species; and (2) setae z5 in P. ainscoughi less than half as long as s5, and also shorter than adjacent setae in j-series (Figures 4F, 6A), vs. setae z5 only slightly shorter than s5 and slightly longer than adjacent j-series setae in the new species (Figures 2, 4A).
Despite our inability to study all three Iranian deutonymphs in person, we are convinced that they are conspecific. Detailed illustrations of the deutonymph examined by Babaeian (2016) allowed inclusion of data on this specimen for most character systems examined. We have less detail on the specimen collected by Hajizadeh et al. (2021) but a few photographs show that this specimen also has four pairs of setae on the ventral shield, sub-equal marginal setae on the lateral soft cuticle, setae s1 off the dorsal shield, z5 as long as the adjacent setae in the j-series, and setae st1 markedly longer than the other sternal setae. We therefore revise the identification of these specimens from P. punctatus to P. ahangarani n. sp.
Protodinychus ainscoughi Hu\c tu & Călugăr, 2002
Protodinychus ainscoughi Huțu & Călugăr, 2002: 227.
Protodinychus sp. Johnston, 1961: 528 (in part).
Protodinychus johnstoni Hirschmann 1979: 57, nomen nudum.
(Figures 4F, 5–6)


This species was described based on adults and deutonymphs collected from beaver nests in Ontario, Canada. Prior to this, Johnston (1961) reported on two collections of Protodinychus from Ontario, Canada. He figured a female from one of those collections, and made a comment on a protonymph that may have been part of the same collection (from debris in a beaver lodge). The second collection involved deutonymphs phoretic on beaver nest beetles. Conspecificity of this material with P. ainscoughi was rejected by Huțu and Călugăr (2002) because Johnston (1961) described (not figured) a protonymph as having four dorsal shields, while Huțu & Călugăr note only two shields in their specimens (Huțu and Călugăr 2002). We have not been able to recover any of the material of the first collection studied by Johnston (1961) (adults and nymphs) in the OSAL collection, but did recover six of the deutonymphs from Leptonillus validus, presumably from the second collection. These specimens match the description by Huțu and Călugăr (2002), and are therefore provisionally considered conspecific. The status of the material in the first collection is considered unknown until actual specimens can be recovered.
In a final complication, Hirschmann (1979a) designated Johnston's specimens as P. johnstoni Hirschmann, 1979, based exclusively on Johnston's (1961) drawings of a female. This is considered a nomen nudum because according to article 13.1.1 of the Code for Zoological Nomenclature (1999) in order for a name to be available, it needs to be accompanied by a description or definition that states in words the characters that differentiate the taxon, and neither Hirschmann or Johnston provided such data.
In the original description of the species, setae j1 in the deutonymph were listed as inserted on soft cuticle. However, examination of the specimens associated with L. validus indicated these setae are inserted on a lightly sclerotized vertex plate (Figure 5B). Other additions to the earlier description of the deutonymph include: (1) a pore in the median dorsal region of the gnathotectum; (2) tibia II in this species has only eight setae, including only one pl seta (2, 1/1, 2/1, 1), instead of the nine setae in P. ahangarani n. sp. and P. punctatus (2, 1/1, 2/1, 2) (Evans 1972).
Material examined
Six deutonymphs ex Leptinillus validus (Col.: Leiodidae) collected in debris of a beaver lodge in Provoking Lake, Algonquin Provincial Park, Ontario, Canada (45.5581° N, 78.4919° W), 30 July 1956, coll. D.M. Wood and deposited in the Acarology Laboratory of the Ohio State University (OSAL 013579, 043188-043189, 444423-044425).
Key to deutonymphs of Protodinychus
1. Ventral shield with 2 pairs of setae
...... 2
—Ventral shield with 4 pairs of setae
...... 3
2. Dorsal shield with 22 pairs of setae, seta s1 on the shield; st4 off sternal shield
...... P. punctatus
— Dorsal shield with 20 pairs of setae, seta s1 off the shield; st4 on sternal shield
...... P. evansi
3. Setae s1 on dorsal shield and short, as long as z1; dorsolateral soft cuticle setae variable in length; setae z5 less than half the length of s5 and shorter than adjacent j-series setae (Figure 4F)
...... P. ainscoughi
— Setae s1 off dorsal shield and long, as long as j1; dorsolateral soft cuticle setae long, subequal in length; setae z5 slightly shorter than s5, slightly longer than adjacent j-series setae (Figure 4A)
...... P. ahangarani n. sp.
Discussion
The current findings suggest that the geographic range of the genus Protodinychus is broader than currently known, covering the temperate zones of the western Palearctic and eastern North America. Previous records for the genus suggest a very large range for a single species, P. punctatus but our observations suggest an alternative hypothesis with a number of similar, and possibly geographically restricted, species. This would imply some undiscovered diversity, and re-examination of records from areas far away from the type locality in England might be indicated. What we cannot explain is why there are so few records of Protodinychus (and Thinozercon) given that these are quite distinctive mites. Is the currently established geographic range a true reflection of the range of these genera (suggesting relict distributions) or will we see many more collections e.g. from areas in the eastern Palearctic and western North America?
Mites in this genus may also have a broader ecological range than previously assumed. Existing records of P. punctatus are mostly from nutrient poor grasslands (e.g. Athias-Binche and Evans 1981; Siepel et al. 2018), but P. ainscoughi has only been found in beaver lodges (high nutrient environment) while the Iranian records are from litter in fairly rich and diverse forest environments (similar to P. evansi). For a small number of records, this suggests an impressive ecological breath, a situation not unlike that found in e.g. Opilioacaridae (Vázquez and Klompen 2015). These observations are also consistent with a hypothesis that Protodinychidae is a relict group.
Acknowledgments
The authors wish to thank Yazdanfar Ahangaran for his valuable help in sampling the material used in this study; Alireza Saboori and Jalil Hajizadeh for sharing information about the previously collected specimens from Kheyroud-Kenar Forest and Anzali City, respectively, which were reported as P. punctatus, and Bruce Halliday for help on nomenclatural issues concerning P. johnstoni. Financial support to HK provided in part by National Science Foundation grant DEB 2017439.
References
- Athias-Binche F., Evans G.O. 1981. Observations on the genus Protodinychus Evans, 1957 (Acari: Mesostigmata) with descriptions of the male and phoretic deuteronymph. Proc. R. Irish Acad. (Dublin), 81B: 25-36.
- Babaeian E. 2016. Identification of Uropodina mites in Kheyroud-Kenar forest, Mazandaran province [Ph. D. thesis]. Tehran: University of Tehran.
- Chaudhury S., Roy I., Podder S., Gupta S.K., Saha G.K. 2005. Diversity of synanthropic mites in Kolkata Metropolis, India. Rec. zool. Surv. India, 104: 151-159.
- Evans G.O. 1957. An introduction to the British Mesostigmata (Acarina) with keys to families and genera. Zool. J. Linn. Soc., 43: 203-259. https://doi.org/10.1111/j.1096-3642.1957.tb01552.x
- Evans G.O. 1963. Some observations on the chaetotaxy of the pedipalps on the Mesostigmata (Acari). Ann. Mag. Nat. Hist., 6: 513-527. https://doi.org/10.1080/00222936308651393
- Evans G.O. 1972. Leg chaetotaxy and the classification of the Uropodina (Acari: Mesostigmata). J. Zool., Lond., 167: 193-206. https://doi.org/10.1111/j.1469-7998.1972.tb01729.x
- Hajizadeh J., Moammadi L., Mirhaghparast S.K. 2021. Introduction of two rare mites (Acari) species from northern Iran. J. Biol. Stud., 4: 73-81.
- Hirschmann W. 1979a. Gangsystematik der Parasitiformes. Teil 338. Bestimmbare Uropodiden-Arten der Erde (ca. 1200 Arten), geordnet nach dem Gangsystem Hirschmann 1979 und nach Adulten-Gruppen (Stadien, Heimatländer, Synonym, Literatur). Acarologie, 26: 15-57.
- Hirschmann W. 1979b. Stadiensystematik der Parasitiformes. Teil 1. Stadienfamilien und Stadiengattungen der Atrichopygidiina, erstellt im Vergleich zum Gangsystem Hirschmann 1979. Acarologie, 26: 57-68.
- Huțu M., Călugăr A. 2002. Zwei neue Protodinychus-Arten (Anactinotrichida: Uropodina: Protodinychidae). Abh. Ber. Naturkundemus. Görlitz, 74: 219-236.
- Johnston D.E. 1961. A review of the lower uropodoid mites (former Thinozerconoidea, Protodinychoidea, and Trachytoidea) with notes on the classification of the Uropodina. Acarologia, 3: 522-545.
- Karg W. 1989. Acari (Acarina), Milben, Unterordnung Parasitiformes (Anactinochaeta), Uropodina Kramer, Schildkrötenmilben. Jena: Gustav Fischer Verlag. pp. 1-203.
- Klompen H. 2000. A preliminary assessment of the utility of elongation factor-1alpha in elucidating relationships among basal Mesostigmata. Exp. Appl. Acarol., 24: 805-820. https://doi.org/10.1023/A:1006432017638
- Klompen H., Lekveishvili M.G., Black W.C., IV. 2007. Phylogeny of Parasitiform mites (Acari) based on rRNA. Mol. Phylogen. Evol., 43: 936-951. https://doi.org/10.1016/j.ympev.2006.10.024
- Kontschán J. 2014. Uropodina mites (Acari: Mesostigmata) of Transylvania (Romania). Budapest: Ad Librum.
- Kontschán J. 2015. First record of the family Protodinychidae Evans, 1957 (Acari: Mesostigmata: Uropodina) from Hungary. Ecol. Montenegrina, 2: 283-288. https://doi.org/10.37828/em.2015.2.33
- Krantz G.W., Ainscough B.D. 1990. Acarina: Mesostigmata (Gamasida). In: Dindal D.L. (Ed). Soil Biology Guide. New York: John Wiley & Sons, Inc. p. 583-665.
- Lindquist E.E., Krantz G.W., Walter D.E. 2009. Order Mesostigmata. In: Krantz G.W., Walter D.E. (Eds). A manual of acarology. Lubbock, TX: Texas Tech University Press. p. 124-232.
- Napierala A., Bloszyk J. 2013. Unstable microhabitats (merocenoses) as specific habitats of Uropodina mites (Acari: Mesostigmata). Exp. Appl. Acarol., 60: 163-180. https://doi.org/10.1007/s10493-013-9659-9
- Norton R.A., Kethley J.B., Johnston D.E., OConnor B.M. 1993. Phylogenetic perspectives on genetic systems and reproductive modes of mites. In: Wrensch D.L., Ebbert M.A. (Eds). Evolution and diversity of sex ratio in insects and mites. New York: Chapman & Hall. p. 8-99. https://doi.org/10.1007/978-1-4684-1402-8_2
- Pugh P.J.A., King P.E., Fordy M.R. 1987. Structural Features Associated with Respiration in Some Intertidal Uropodina (Acarina, Mesostigmata). J. Zool., 211: 107-120. https://doi.org/10.1111/j.1469-7998.1987.tb07456.x
- Salmane I., Spungis V. 2015. Factors influencing Mesostigmata mites (Acari, Parasitiformes) in the alkaline fen habitats. Proc. Latv. Acad. Sci., Sect. B Nat. Exact Appl. Sci., 69: 50-56. https://doi.org/10.1515/prolas-2015-0006
- Siepel H., Cremers H.J., Dimmers W.J., Loomans A., Vierbergen B. 2018. Checklist of the mesostigmatic mites of the Netherlands (Acari: Mesostigmata). Ned. Faun. Meded., 51: 115-188.
- Vázquez M.M., Klompen H. 2015. The family Opilioacaridae (Parasitiformes: Opilioacarida) in Mexico, description of two new species and notes on biology and geographical distribution. Zootaxa, 3957: 535-552. https://doi.org/10.11646/zootaxa.3957.5.3
- Wisniewski J., Hirschmann W. 1993. Gangsystematik der Parasitiformes. Teil 548, Katalog der Ganggattungen, Untergattungen, Gruppen and Arten der Uropodiden der Erde (Taxonomie, Literatur, Grösse, Verbreitung, Vorkommen. Acarologie, 40: 1-220.



2022-03-04
Date accepted:
2022-05-16
Date published:
2022-06-09
Edited by:
Faraji, Farid

This work is licensed under a Creative Commons Attribution 4.0 International License
2022 Kazemi, Shahrooz and Klompen, Hans
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)