



A new water mite species of the genus Teutonia Koenike, 1889 from Corsica, France, based on morphological data and DNA barcodes (Acari, Hydrachnidia, Teutoniidae)
Pešić, Vladimir1 and Smit, Harry2
1✉ Department of Biology, University of Montenegro, Cetinjski put b.b., 81000 Podgorica, Montenegro.
2Naturalis Biodiversity Center, P.O. Box 9517, 2300 RA Leiden, the Netherlands.
2022 - Volume: 62 Issue: 2 pages: 465-473
https://doi.org/10.24349/8n5y-mw7lZooBank LSID: F83B5FED-0C63-41BC-A2F5-A331E03CD95E
Original research
Keywords
Abstract
Introduction
The family Teutoniidae Koenike, 1910 comprises of two genera, the monotypic Limnolegeria Motaş, 1928, which was for a long time considered as a representative of the Anisitsiellidae (see Cook 1974), and the widely distributed Teutonia Koenike, 1889. The latter genus, recorded from the Holarctic, comprises of two subgenera, the nominate subgenus and Subteutonia Habeeb, 1958 known only from Canada (Smit 2020).
Currently, two species of the genus Teutonia are known from the Western Palaearctic (Di Sabatino et al. 2010), i.e., the widely distributed T. cometes (Koch, 1837), inhabiting cold montane lakes and pools of shaded streams (Di Sabatino et al. 2010), and T. subalpina Thor, 1897. The latter, little known species is restricted to the far north of Europe (alpine and subalpine coniferous forests of Norway and Sweden) (Di Sabatino et al. 2010).
In this paper we used morphological data and DNA barcode of the mitochondrial cytochrome c oxidase subunit I (COI) gene sequence to describe one new species of the genus Teutonia from Corsica.
Material and methods
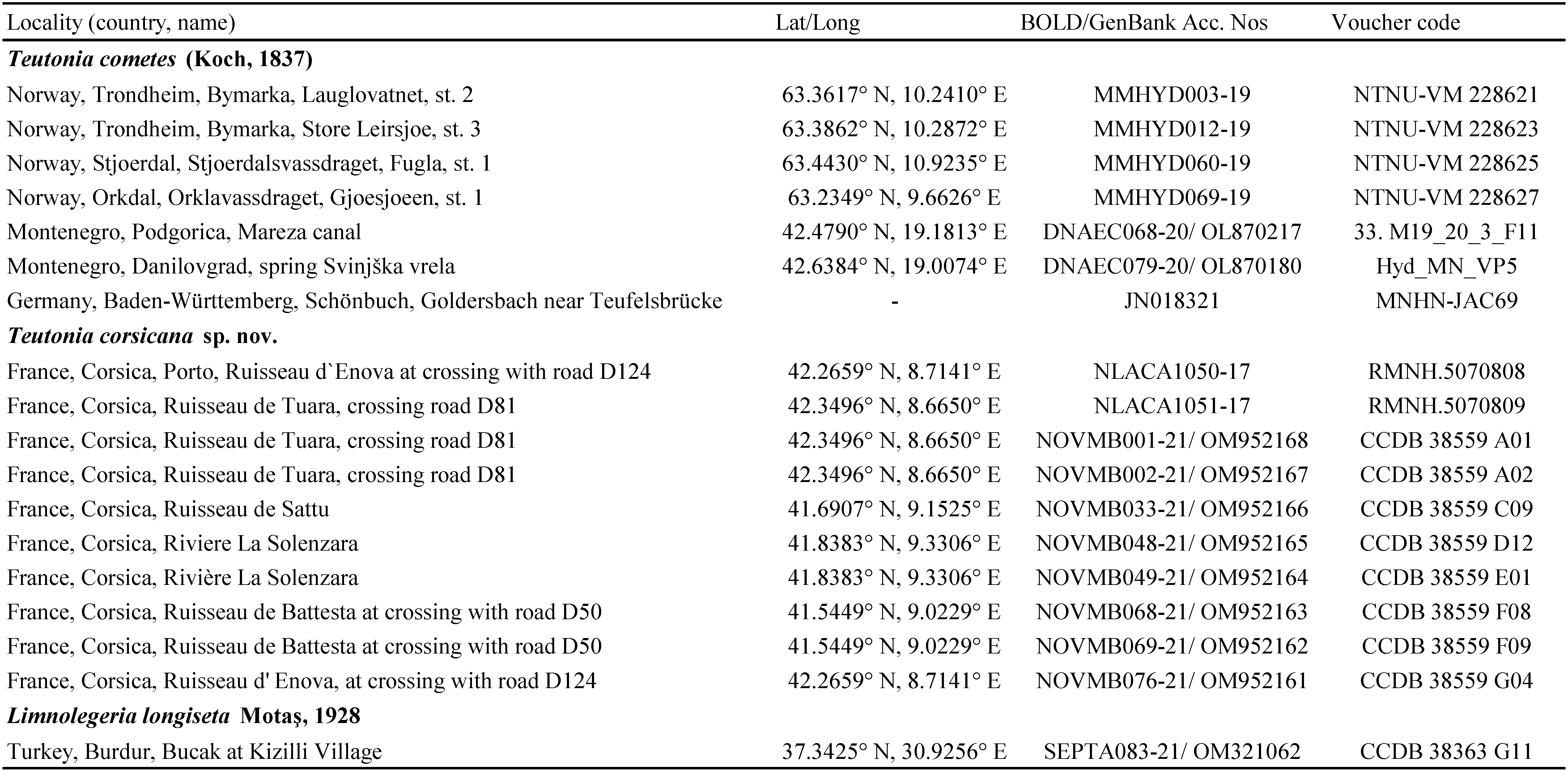
Water mites were collected by hand netting by the junior author, sorted live in the field, and immediately preserved in 96% ethanol for the purpose of the molecular analyses. A part of the material was also preserved in Koenike-fluid. After DNA extraction, two voucher specimens were dissected and slide mounted in Faure's medium. Holotype and paratypes of the new species are deposited in Naturalis Biodiversity Center in Leiden (RMNH). The DNA sequences prepared in the course of this study were deposited in the Barcode of Life Data System (BOLD) and GenBank (Table 1).

All measurements are in µm. Morphological nomenclature follows Gerecke et al. (2016). The length of genital flaps and acetabula were measured on both sides, and therefore their dimensions are given as a range. The photograph of ejaculatory complex was made using a camera on a Samsung Galaxy smartphone. The following abbreviations are used: Ac – acetabula; a.s.l. – above sea level; Cx-I-IV – first-fourth coxae; dL – dorsal length; H – height; I-IV -L-1-6 – first-sixth segments of first- fourth legs; L – length; P-1-P-5 – palp segment 1-5; RMNH – Naturalis Biodiversity Center, Leiden; vL – ventral length; W – width.
Molecular analyses were conducted at the Canadian Centre for DNA Barcoding (Guelph, Ontario, Canada (CCDB; http://ccdb.ca/ ![]() )), where the specimens were sequenced for the barcode region of COI using standard invertebrate DNA extraction, amplification and sequencing protocols (Ivanova et al. 2007; Ivanova and Grainger 2007a, b). In CCDB the chromatograms were assembled into consensus sequences for each specimen and uploaded to the barcode of life database (BOLD; https://www.boldsystems.org/
)), where the specimens were sequenced for the barcode region of COI using standard invertebrate DNA extraction, amplification and sequencing protocols (Ivanova et al. 2007; Ivanova and Grainger 2007a, b). In CCDB the chromatograms were assembled into consensus sequences for each specimen and uploaded to the barcode of life database (BOLD; https://www.boldsystems.org/ ![]() ). The sample identifiers in BOLD are given for each barcoded specimen.
). The sample identifiers in BOLD are given for each barcoded specimen.
The final dataset for the phylogenetic analysis includes 17 COI sequences representing Teutonia cometes (n=7), and T. corsicana sp. nov. (n=10). Limnolegeria longiseta Motaş, 1928 (BOLD/GenBank SEPTA083-21/OM321062) was used as outgroup (Table 1).
Sequence comparisons were performed using MUSCLE alignment (Edgar 2004). Intra- and interspecific genetic distances were calculated based on the Kimura 2-parameter model (K2P; Kimura 1980), using MEGA-X (Kumar et al. 2018). MEGA-X software was used to calculate Neighbour-Joining (NJ) trees based on K2P distances (standard for barcoding studies) and pairwise deletion of missing data. The support for tree branches was calculated by the non-parametric bootstrap method (Felsenstein 1985) with 1000 replicates. Codon positions included were 1st+2nd+3rd+Noncoding. All ambiguous positions were removed for each sequence pair.
In order to assess the genetic differentiation of species we used the online ASAP (assemble species by automatic partitioning) version (https://bioinfo.mnhn.fr/abi/public/asap/asapweb.html ![]() ) with default settings and the K2P distance model. The latter procedure was designated to a list of partitions of species hypotheses using genetic distances, calculated between DNA sequences and ranked by their ASAP-scores: the lower the score, the better the partition (Puillandre et al. 2021).
) with default settings and the K2P distance model. The latter procedure was designated to a list of partitions of species hypotheses using genetic distances, calculated between DNA sequences and ranked by their ASAP-scores: the lower the score, the better the partition (Puillandre et al. 2021).
Results and discussion
Species delimitation using DNA-barcodes
The final alignment for species delimitation using COI sequence data included 17 Teutonia specimens listed in Table 1 and one outgroup, Limnolegeria longiseta from Turkey. The final alignment consisted of 837 nucleotide positions. The NJ tree is presented in Figure 1.

The sequences retrieved from the Teutonia specimens from Corsica appeared as a sister clade of the clade that encompasses specimens of T. cometes from Norway, Germany and Montenegro (Figure 1). The average K2P genetic distance between T. corsicana sp. nov. from Corsica and T. cometes was estimated 18.7±1.9%. Both species possesses a low intraspecific distance, 0.15±0.07% K2P for T. corsicana sp. nov. and 0.36±0.15% K2P for T. cometes.
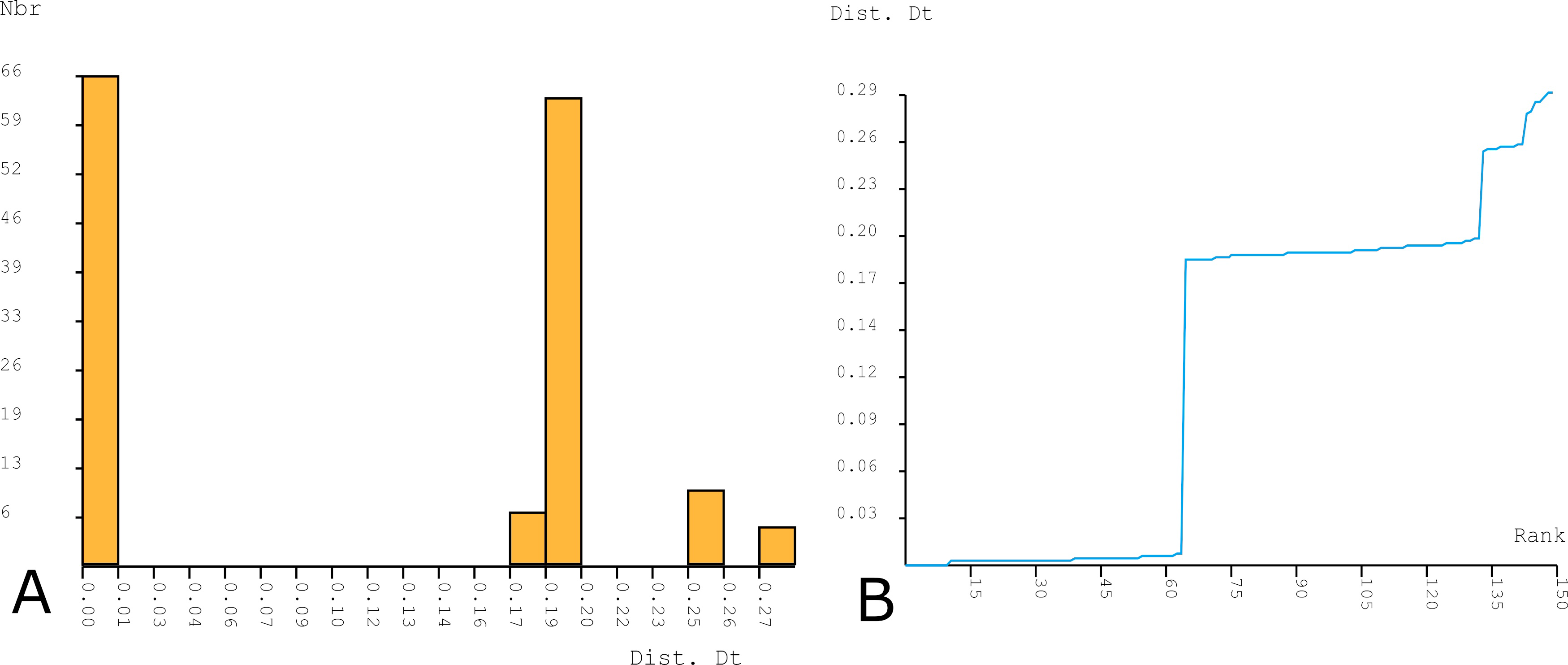
The ASAP method observed a barcode gap of about 1-18% K2P distances (Figure 2A-B). The applied ASAP procedure identified two MOTUs (hypothetical species) corresponding to T. cometes and T. corsicana sp. nov. at the threshold distance of 9.51% (K2P) which has the best ASAP-score (1.00) within the available molecular data.
Systematics
Family Teutoniidae Koenike, 1910
Diagnosis — Smit 2020: 268.
Genus Teutonia Koenike, 1889
Diagnosis — Smit 2020: 269.
Teutonia corsicana sp. nov.
ZOOBANK: 5877B3D9-E264-4102-A442-3356BCE46C10 ![]()
Figures 3-4
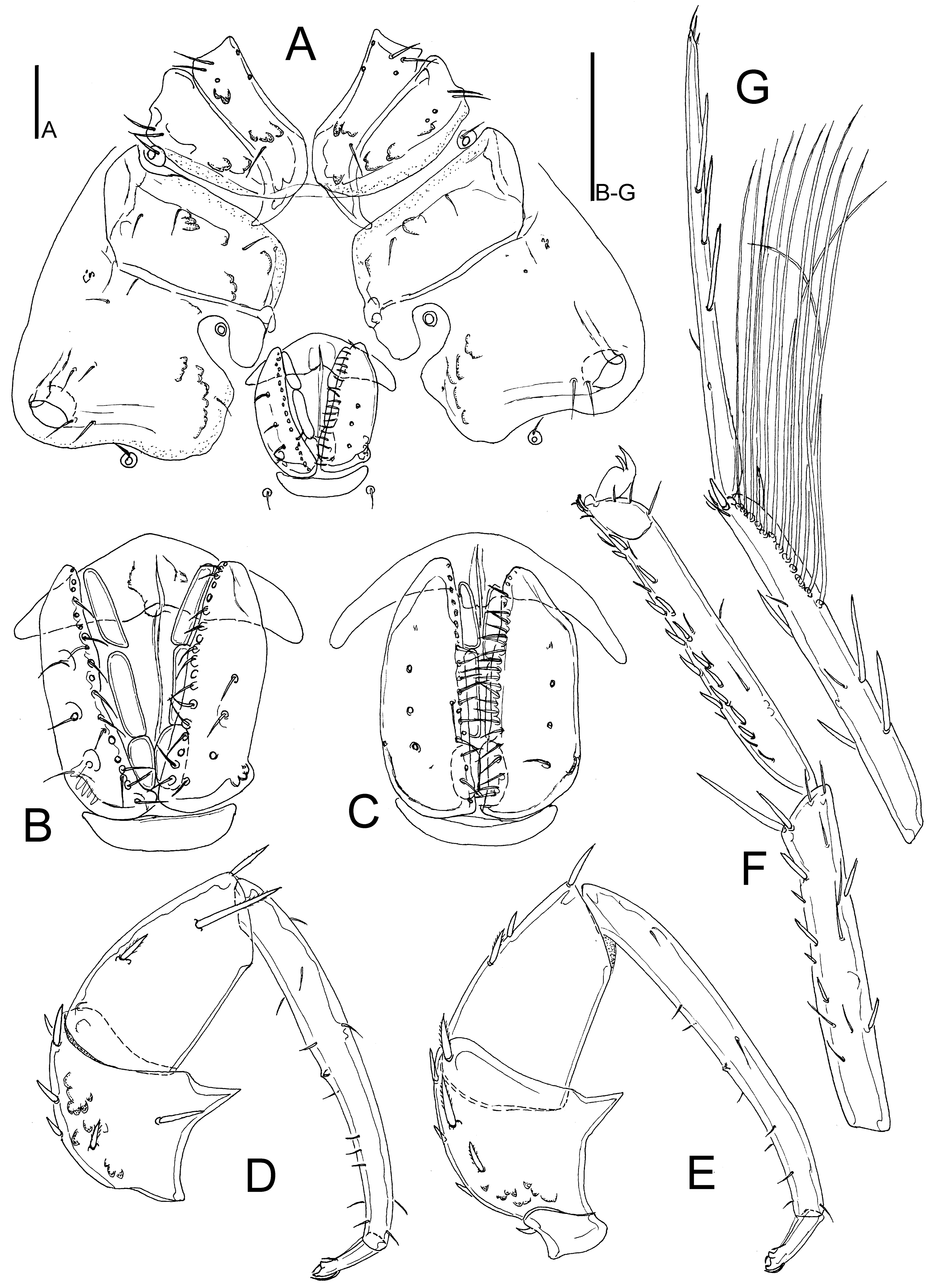
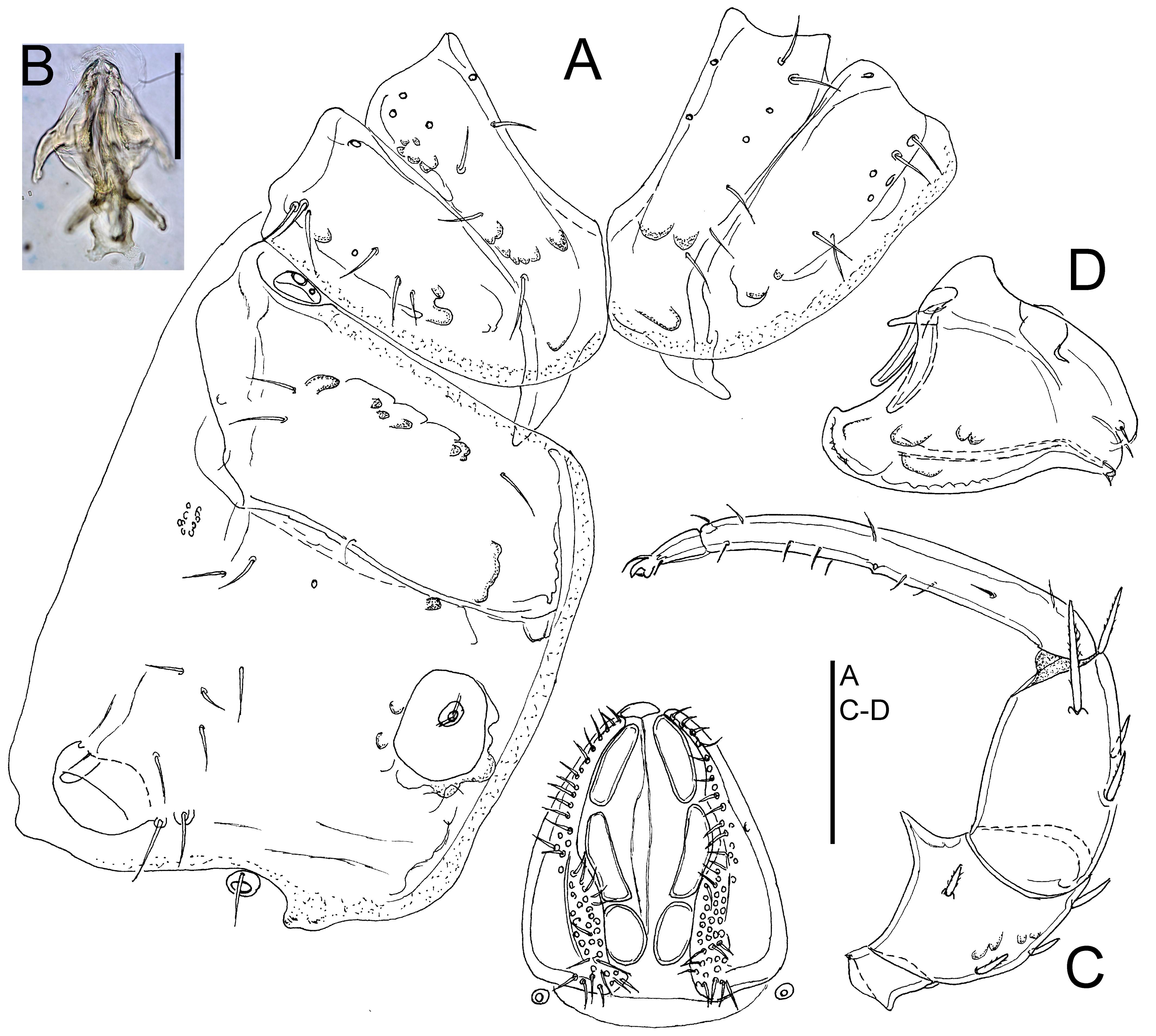
Material examined
Holotype ♀, France, Corsica, Ruisseau de Tuara, crossing road D81, 42.3496° N, 8.66497° E, 238 m a.s.l., 14 Apr. 2015, leg. Smit (sequenced [voucher code: CCDB 38559 A01; BOLD/GenBank Nos.: NOVMB001-21/ OM952168]), dissected and slide mounted. Paratype: 1 ♀, same place and data as holotype, sequenced (Table 1); 1♂, 1♀, Ruisseau de Sattu, Corsica, 41.6907° N, 9.1525° E, 370 m a.s.l., 10 Apr. 2015, leg. Smit, 1♂ sequenced (Table 1); 1♂, 1♀, Rivière La Solenzara, Corsica, 41.8383° N, 9.33055° E, 90 m a.s.l., 7 Apr. 2015, leg. Smit, 1♂, 1♀ sequenced (Table 1); 2♂, Ruisseau de Battesta at crossing with road D50, Corsica, 41.5449° N, 9.02292° E, 220 m a.s.l., 8 Apr. 2015, leg. Smit, 2♂ sequenced (Table 1), 1♂ (voucher code: CCDB 38559 F09) dissected and slide mounted; 2♀, Ruisseau de Enova, at crossing with road D124, Corsica, 42.2659° N, 8.71405° E, 105 m a.s.l., 13 Apr. 2015, leg. Smit, 1♀ sequenced (Table 1).
Other material
France, Corsica: Ruisseau de Battesta, at crossing with road D50, 41.54487° N, 9.02292° E, 220 m a.s.l., 8 Apr. 2015, 3♀, 1 deutonymph; Ruisseau de Campaghiu, Porto, 42.24128° N, 8.73240° E, 188 m a.s.l., 11 Apr. 2015, 1♀, 1 deutonymph; Ruisseau d'Aitone, at crossing with road D84, 42.26365° N, 8.84597° E, 1115 m a.s.l., 12 Apr. 2015, 1♀; Ruisseau de Casa Infurcata, at crossing with road D124, 42.25600° N, 8.76513° E, 217 m a.s.l., 13 Apr. 2015, 1♀; Ruisseau d'Enova, at crossing with road D124, 42.26590° N, 8.71405° E, 105 m a.s.l., 13 Apr. 2015, 1♂, 5♀, 2 deutonymphs; Rivière Le Fango, at crossing with road D81, 42.41503° N, 8.68300° E, 6 m a.s.l., 14 Apr. 2015, 1♀, 4 deutonymphs; Ruisseau de Tuara, at crossing with road D81, 42.34958° N, 8.66497° E, 230 m a.s.l., 14 Apr. 2015, 3♀, 4 deutonymphs; tributary of Ruisseau de Canne, at crossing with road D81, 42.38307° N, 8.69903° E, 225 m a.s.l., 14 Apr. 2015, 1♀, 2 deutonymphs; Rivière de Tartagine, at Maison Forestière Tartagine, 42.49410° N, 8.99288° E, 706 m a.s.l., 16 Apr. 2015, 1♀; unnamed stream N of Suarella, at crossing with road D18, 42.39033 N, 9.13762° E, 387 m a.s.l., 17 Apr. 2015, 1♀.
Diagnosis
Female gonopore pregenital sclerite partly covered by genital flaps; in both sexes P-2 with broad-based ventrodistal projection, P-3 slender (L/H ratio 1.6-1.7).
Description
General features — Integument finely striated. Coxae medially touching, but not fused, with a few setae. Ac-1-2 rectangular, Ac-3 roundish. P-2 with a generally broad-based ventrodistal projection, P-3 with nearly straight, or slightly convex ventral margin, P-3 lateral seta long, shifted to medial surface, P-4 slender, bearing several fine hair-like setae, mainly in anteroventral part. Legs: claws with claw blade and ventral clawlets; swimming setae restricted to III-/IV-L-3-4, numbers: III-L-3, 3; III-L-4, 8; III-L-5, 8-9; IV-L-3, 3; IV-L-4, 8-9; IV-L-5, 11-12. Male — Glandularium on Cx-IV completely surrounded by coxal sclerotization. Genital field: genital flaps medially with a dense cover of fine setae, pregenital sclerite small, postgenital sclerite as wide as genital field. Female — Glandularium on Cx-IV partly surrounded by coxal sclerite, medially in contact with the perigenital membrane. Genital field: genital flaps with a row of regularly arranged medial setae, pregenital sclerite large, crescent shaped, partly covered by genital flaps (but only slightly covered in Koenike-fixed specimens), gonopore shorter than the genital flaps, postgenital sclerite shorter than pregenital sclerite.
Description — Female (holotype) – Idiosoma L 1190, W 905. Coxal field: L 581, W 878; Cx-III W 563; L Cx-I+II 228, Cx-III+IV 434. Genital flaps L 177-181; pregenital sclerite W 200, postgenital sclerite W 106; L Ac-1-3: 59, 56-60, 39-43. Eggs maximum diameter (n=2) 173-178.
Palp total L 684, dL/H, dL/H ratio: P-1, 33/44, 0.75; P-2, 142/117, 1.21; P-3, 164/102, 1.61; P-4, 289/41, 7.1; P-5, 56/15, 3.8; L ratio P-2/P-4 0.49. Gnathosoma vL 194; chelicera L 256, basal segment L 219, claw L 44, L ratio chelicera basal segment/claw 5.0. Leg segments dL: I-L-2-6: 122, 166, 228, 244, 253; IV-L-1-6: 156, 156, 200, 253, 264, 331.
Male (paratype, CCDB 38559 F09) – Idiosoma L 750, W 588. Coxal field: L 495, W 703; Cx-III W 459; L Cx-I+II 197, Cx-III+IV 380. Genital field L/W 166/147, genital flaps L 156-163, pregenital sclerite W 119; L Ac-1-3: 50, 52-53, 34-38. Ejaculatory complex (Figure 3B) L 192.
Palp total L 581, dL/H, dL/H ratio: P-1, 28/36, 0.77; P-2, 120/96, 1.25; P-3, 142/84, 1.69; P-4, 244/34, 7.2; P-5, 47/14, 3.3; L ratio P-2/P-4 0.49. Gnathosoma vL 165; chelicera L 230, basal segment L 186, claw L 44, L ratio chelicera basal segment/claw 4.3. Leg segments dL: I-L-1-6: 71, 111, 135, 191, 200, 209; IV-L-1-6: 131, 144, 175, 215, 236, 284.
Etymology
Named after the island where the new species was discovered.
Remarks
The female of the new species from Corsica, is easily distinguished from the two other know species of the genus, T. cometes and T. subalpina in the shape of genital field having a pregenital sclerite partly covered by the genital flaps. Moreover, in both sexes T. cometes differs in P-2 generally with a narrow-based ventrodistal projection (but in larger populations, the latter projection appearing broad-based in some specimens, especially in juveniles) and stouter P-3 (L/H ratio 1.35-1.5 vs. 1.6-1.7 in T. corsicana sp. nov.). The high genetic distance between the new species and T. cometes (COI 18.7±1.9 K2P%) suggests a long independent history of the new species. The former records of T. cometes from Corsica (Angelier 1959; Santucci 1971) likely should be assigned to the new species.
The little known T. subalpina can be distinguished in the female by the very long gonopore, anteriorly distinctly extending over the margin of the genital flaps (Di Sabatino et al. 2010). The taxonomic status of T. subalpina is still unclear, and according to Di Sabatino et al. (2010), it is possible that the original description of the latter species was based on freshly hatched, unsclerotized, misshapen adults of T. cometes.
Distribution
France (Corsica).
Key to the European species of Teutonia Koenike, 1889
1. Female gonopore very long, only the posterior 2/3 flanked by Ac and genital flaps; ventral projection of P-2 in both sexes on a large base, triangular
...... Teutonia subalpina
— Female gonopore all over it length flanked by Ac and genital flaps; ventral projection of P-2 various
...... 2
2. Female pregenital sclerite not covered by genital flaps (as illustrated in 5A); ventral projection of P-2 in both sexes on a narrow base, peg-like (as illustrated in Figure 5B); P-3 stout, L/H ratio 1.35-1.5
...... Teutonia cometes
— Female pregenital sclerite partly covered by genital flaps; ventral projection of P-2 in both sexes on a large base, triangular; P-3 slender, L/H ratio 1.6-1.7
...... Teutonia corsicana sp. nov.

Acknowledgements
We are indebted to Elisabeth Stur (Trondheim, Norway) and Reinhard Gerecke (Tübingen, Germany) for permission to use some unpublished sequences from Norway (Project ''Water M&M (University of Trondheim)''. Special thanks to Milica Jovanović and Ana Manović (University of Podgorica) for their excellent routine laboratory processing work. HS would like to thank Truus van der Pal (Alkmaar) for her assistance with the field work on Corsica. This study is part of the ''DNA-Eco'' scientific project, supported by a grant of the Montenegrin Ministry of Science. We thank Joanna Mąkol (Wrocław), Hiroshi Abé (Fujisawa) and one anonymous reviewer, whose constructive comments greatly improved this work.
References
- Angelier E. 1959. Acariens (Hydrachnellae et Porohalacaridae) des eaux superficielles. In: Angelier E. et collaborateurs, Hydrobiologie de la Corse. Vie et Milieu, Suppl., 8: 64-148.
- Cook D.R. 1974. Water mite genera and subgenera. Mem. Am. Entomol. Inst., 21: 1-860.
- Di Sabatino A., Gerecke R., Gledhill T., Smit H. 2010. Chelicerata: Acari II. In: Gerecke R. (ed.) Chelicerata: Araneae, Acari I. Süßwasserfauna von Mitteleuropa, 7, 2-2: 1-134. München: Elsevier Spektrum Akademischer Verlag. https://doi.org/10.1007/978-3-8274-2266-8_1
- Edgar R.C. 2004. MUSCLE: multiple sequence alignment with high accuracy and high 679 throughput. Nucleic acids Res 32(5): 1792-1797. https://doi.org/10.1093/nar/gkh340
- Felsenstein J. 1985. Confidence limits on phylogenies: An approach using the bootstrap. Evolution, 39: 783-791. https://doi.org/10.1111/j.1558-5646.1985.tb00420.x
- Gerecke R., Gledhill T., Pešić V., Smit H. 2016. Chelicerata: Acari III. In: Gerecke R. (ed.) Süßwasserfauna von Mitteleuropa, Bd. 7/2-3. Springer-Verlag Berlin, Heidelberg, pp. 1-429. https://doi.org/10.1007/978-3-8274-2689-5
- Ivanova N.V., de Waard J.R., Hebert P.D.N. 2007. CCDB protocols, glass fiber plate DNA extraction. ccdb.ca/site/wp-content/uploads/2016/09/CCDB_DNA_Extraction.pdf

- Ivanova N.V., Grainger C.M. 2007a. CCDB protocols, COI amplification. Available at: ccdb.ca/site/wp-content/uploads/2016/09/CCDB_Amplification.pdf
- Ivanova N.V., Grainger C.M. 2007b. CCDB protocols, sequencing. Available at: ccdb.ca/site/wp-content/uploads/2016/09/CCDB_Sequencing.pdf
- Kimura M. 1980. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol., 16: 111-120. https://doi.org/10.1007/BF01731581
- Kumar S., Stecher G., Li M., Knyaz C., Tamura K. 2018. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol., 35: 1547-1549. https://doi.org/10.1093/molbev/msy096
- Puillandre N., Brouillet S., Achaz G. 2021. ASAP: assemble species by automatic partitioning. Mol. Ecol. Resour., 21:609-620. https://doi.org/10.1111/1755-0998.13281
- Santucci J. 1971. Contribution à l'étude de la répartition des Hydracariens (Hydrachnellae) des eaux superficielles d'un torrent de Corse - Le Porto. Ann. Fac. Sci. Marseille, 45: 81-99.
- Smit H. 2020. Water mites of the world with keys to the families, subfamilies, genera and subgenera (Acari: Hydrachnidia). Monogr. Nederl. Entom. Veren., 12: 1-774.



2022-03-11
Date accepted:
2022-05-05
Date published:
2022-05-09
Edited by:
Mąkol, Joanna

This work is licensed under a Creative Commons Attribution 4.0 International License
2022 Pešić, Vladimir and Smit, Harry
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)