



Water mites of the genus Lebertia Neuman, 1880 from the eastern Himalayas (Acari: Hydrachnidia: Lebertiidae)
Gerecke, Reinhard1 and Smit, Harry2
1Biesingerstraße 11, DE 72070 Tübingen, Germany & Arachnology, Senckenberg, Senckenberganlage 25, D-60325 Frankfurt, Germany.
2✉ Naturalis Biodiversity Center, P.O. Box 9517, 2300 RA Leiden, the Netherlands.
2022 - Volume: 62 Issue: 2 pages: 302-316
https://doi.org/10.24349/esot-nc22ZooBank LSID: 52668A17-CAAF-437A-864C-A2883EF87409
Original research
Keywords
Abstract
Introduction
One of the largest mountain ranges in the world are the Himalayas, stretching from Pakistan in the west to the Indian state of Arunchal Pradesh in the east over a distance of 2,400 km.
The knowledge of water mites of this mountain range is very poor, and many studies are restricted to the foothills of the Himalayas. Water mites of the Himalayan range (including the foothills) were treated by Walter (1928), Lundblad (1934), Panesar and Gerecke (1994), Panesar (2004), Pešić and Smit (2007), Kumar et al. (2007a, b), Pešić and Panesar (2008) and Pešić and Gerecke (2008) and Pešić et al. (2012; 2019a, b; 2020a, b).
The identity of the only species of the water mite genus Lebertia so far reported from the Himalayas, L. orientalis Walter, 1928, cannot be elucidated any more. It was described from Himachal Pradesh State (Kangra Valley, Upper Dharamsala, Bhagsu-nath) after a single deutonymph. At this developmental stage, Lebertia specimens do not show the character states essential for morphological species recognition in this genus, and L. orientalis must therefore be considered a nomen dubium. In addition, from the whole Indian subcontinent there is only one further record of a species of the genus: L. carmamaya Cook, 1967, a representative of the subgenus Pilolebertia described from Tamil Nadu State in the south of India (Cook 1967).
Material collected by Piet Veel (Heemstede), Rajan Khatri (Kathmandu), Wim Klein and Oscar Vorst (RMNH) in 2016 shows that Lebertia species are highly abundant in the streams of Nepal and Bhutan. This study presents the taxonomic results of this collecting trip.
Material and methods
All material was collected with a dip net, sorted in the field and fixed in Koenike's fluid. Holotypes and paratypes from Nepal are deposited in the Naturalis Biodiversity Center, Leiden (RMNH), all material from Bhutan is deposited in the National Biodiversity Centre, Thimphu (NBC), a part of the paratypes and the non-type material will be lodged as a long-term loan in Naturalis Biodiversity Center. The following further abbreviations are used (for a detailed illustration of abbreviations used in Lebertia, see Gerecke 2009, Figure 1) : A – P-3, distance between ventrodistal and mediodistal seta; Ac – acetabula; a.s.l. – above sea level; B – P-3, distance between dorsodistal and mediodistal seta; C – P-3, distance between dorsodistal and dorsocentral seta; Cx-I-IV – first to fourth coxae; D – P-3, distance between ventrodistal and dorsocentral seta; dn – deutonymph; E – P-3, distance between dorsocentral and dorsomedial seta; F – P-3, distance between dorsomedial and centrodistal seta; H – height; I-leg – first leg; IV-leg-4-6 – fourth to sixth segments of fourth leg; L – length; mL – mid length; P-1-5 – palp segments 1-5; V1, V3- ventroglandularia 1, 3; W – width. All measurements are in µm. Details given in the descriptions are valid for both sexes if not stated otherwise; differences in measurement values may be deduced from Table 1. Figure 6 gives a selection of characteristic photographs of the collecting sites.
Taxonomy
Family Lebertiidae Thor, 1900
Genus Lebertia Neuman, 1880
Subgenus Lebertia Neuman, 1880
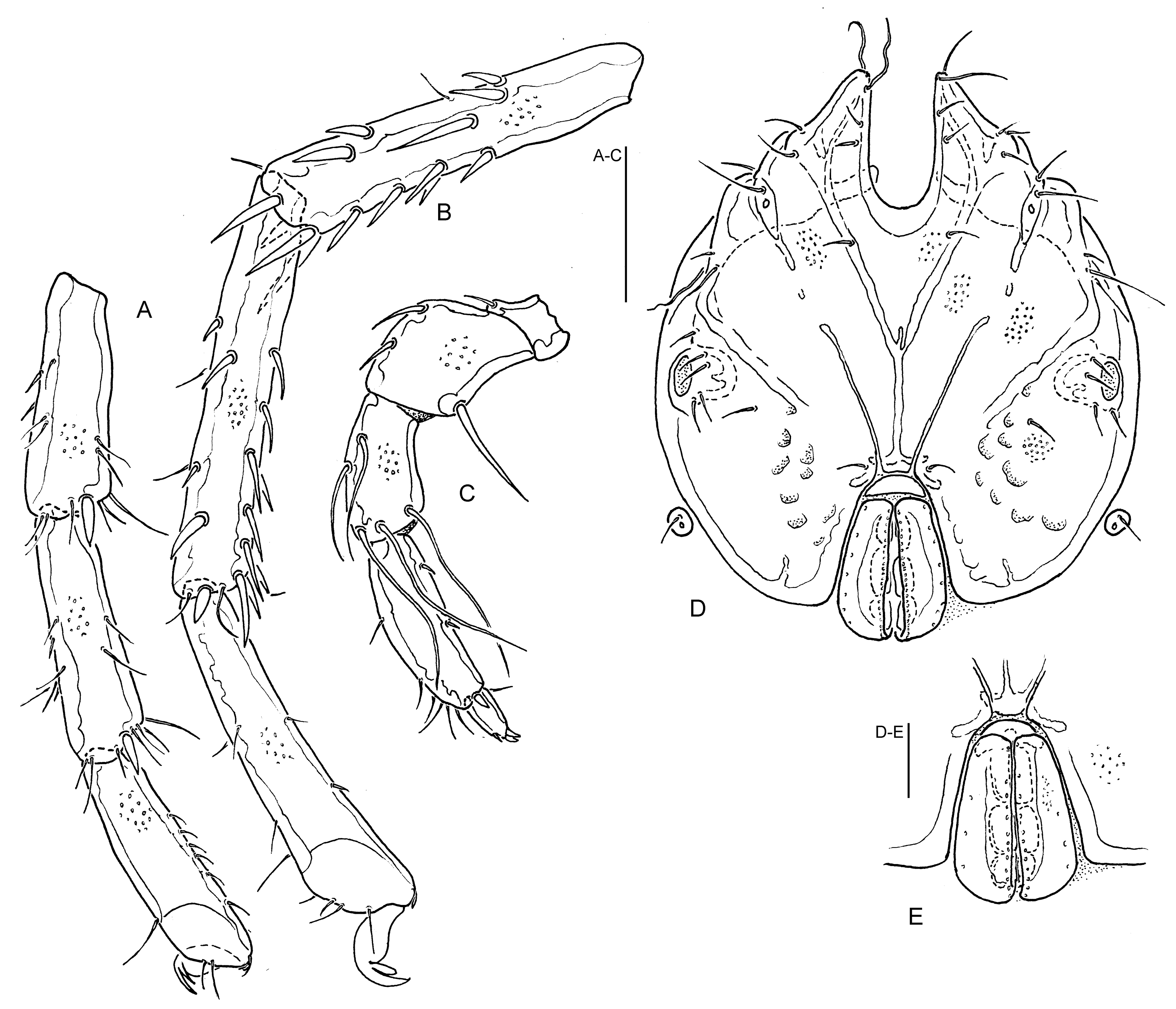
Lebertia (Lebertia) himalayaensis n. sp.
ZOOBANK: 0CD2F98A-2373-4871-97DF-FF4DFFACFA71 ![]()
(Figures 1A-E, 6A)

Type series — Holotype ♂, Bhutan 03, Dumcho, Haa river, pool 3 (Figure 6A), 27°21.883′ N, 89°17.738′ E, 2675 m a.s.l., 07 Aug. 2016, leg. W. Klein, slide mounted (NBC). Paratype ♀, Nepal 03, stream at Jorsalle, 27°46.998′ N, 86°43.062′ E, alt 3101 m, 10 Apr. 2016 leg P.W. Veel, slide mounted (NBC).
Other material examined – Nepal 03, 1 dn slide mounted.
Diagnosis — Integument smooth or with a fine striation; small in dimensions (total L genital field ♂ 123, ♀ 130; total L palp ♂ 234, ♀ 227; L IV-L-4-6, 145; 155-165; 150-155); legs without swimming setae; P-3 mediodistal seta close to dorsodistal seta (A/B 3.4-3.6); P-4 maximum H in distal part, insertions of setae dividing ventral margin into short proximal and distal, and a large central sector(s) (21-24/47-52/28-29 %).
Description — Integument thin and smooth, a fine striation locally visible. Cx-I/-II nearly equal in medial L (ratio 0.9-1.1), posterior margin rather wide (33). Glandularia V3 and setae V1 not fused to coxal shield, V-3 far away from posterolateral margin Cx-IV (Figure 1A). Proportions and setation of distal segments of fourth leg as in Figure 1D; IV-L-6 slightly bowed, with 2-3 fine and short ventral setae; IV-L claw L 35-40. Genital field (Figure 1E) with lateral margins of genital flaps strongly diverging from anterior to posterior end, medial setae (♂: 20, ♀: 9) in a single row. Excretory pore smooth. Gnathosoma with an equally rounded ventral margin (Figure 1B). P-2 relatively slender (L/H 1.49-1.57), ventral seta basally strong, slightly shorter than L of segment (Figure 1C); P-3 rather slender, in basal part not strongly narrowed, dorsal and ventral margin equally diverging from base to tip, with 5 medial setae (2 side by side dorsocentrally, medio-and dorsodistal setae close to each other near distal edge, distoventral seta near segment edge); P-4 club shaped, with dorsal and ventral margins distinctly diverging from base to distal quarter, mediodistal peg seta strong, pointed, ventral setae separated by a wide gap (nearly 50 % segment L), proximoventral setae near proximal, distoventral seta near distal segment end. Measurements: see Table 1.
Etymology — Named for its occurrence in the Himalayas.
Discussion — Lebertia himalayaensis n. sp. differs from many Asian Lebertia s. str. species in the complete absence of swimming setae. Among the four species with the same diagnostic character state, only L. sutschanica Sokolow, 1934 agrees in a smooth integument, for the other three, an integument sculpturation is reported: L. acuta Sokolow, 1934 (reticulation), L. cylindrica Wang & Guo, 2016 and L. pseudomaglioi Wang et al., 2020 (long, resp. short lines). All these species are distinctly major in dimensions (e.g., coxal shield L < 500, genital flap L < 150, P-4 L < 100), and none of these has the ventral setae on P-4 as widely separated as L. himalayaensis n. sp. Among the species known from Europe (Gerecke 2009), in the combination of a smooth integument, legs without swimming setae and a clavate P-4, L. himalayaensis n. sp. is most similar to L. castalia K. Viets, 1925. This species differs from L. himalayaensis n. sp. in the position of P-4 ventral setae (closer to each other, proportions of ventral sectors 2:3:2) and general major dimensions (e.g., palp total L < 320, IV-L-4-6 L < 180). Lebertia fimbriata Thor, 1899 a further European species combining a smooth integument and clavate P-4, differs in the presence of two long distal swimming setae at the penultimate segments of III-L and IV-L. New molecular data indicate that this taxon represents a species group with several so far cryptic species in Europe (unpubl. data). As to judge from the published figures and data, a male from China recently attributed to this species (Wang et al. 2016) is similar to L. fimbriata in shape of idiosoma and palp, but differs from European specimens in proportions and setation of IV-L (stouter, L/H IV-L-4-6, 4.2, 4.7, 4.6, IV-L-5). Thus, it is obviously a representative of a different species.
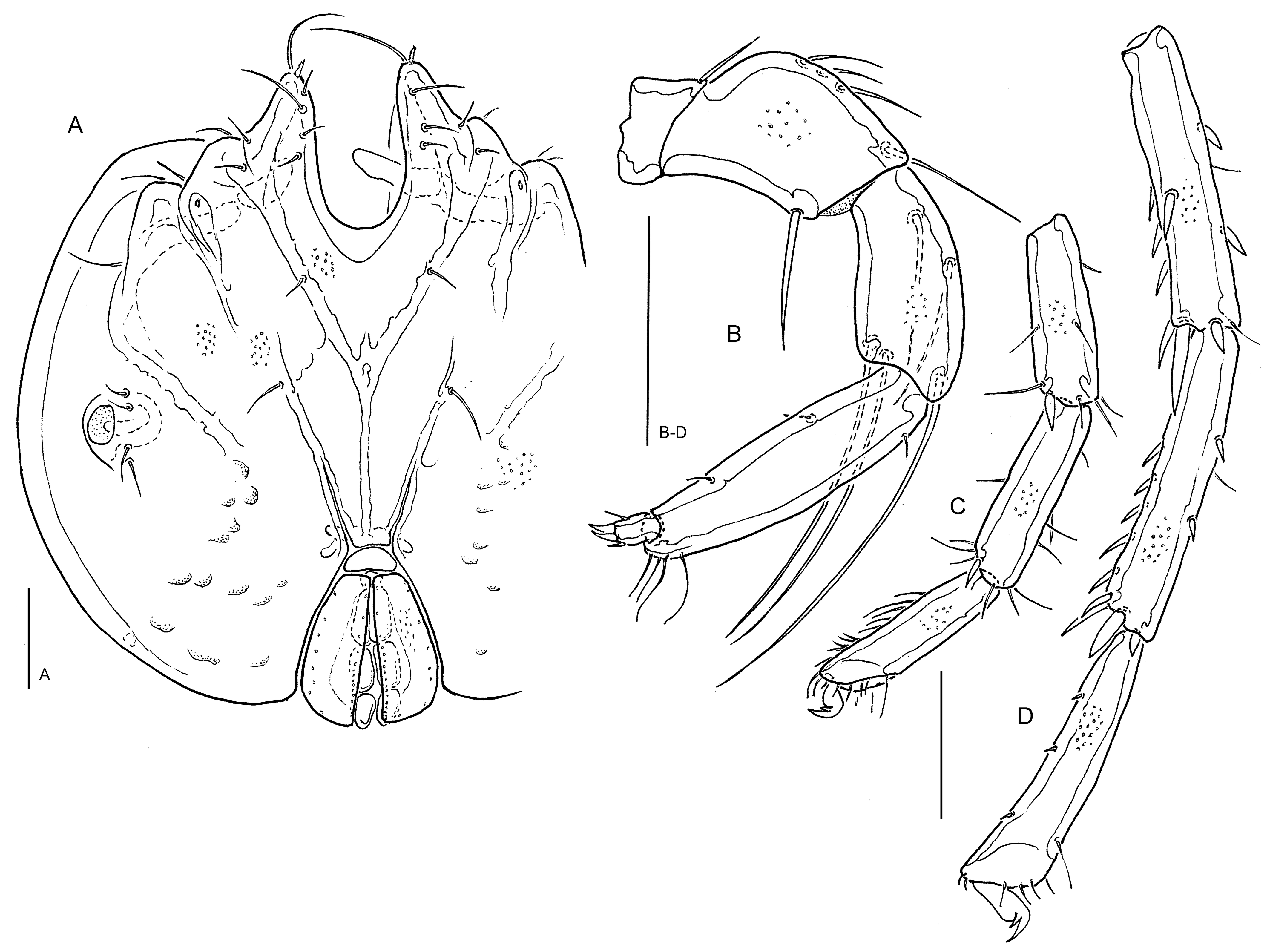
Lebertia (Lebertia) khatrii n. sp.
ZOOBANK: 9FB20554-994E-4E0C-8D7C-D423BD7BFCFB ![]()
(Figures 2A-E, 6B)
Type series — Holotype ♂, Bhutan 01, Bhutan, Royal Botanical Park Lampelri, 27°30.450′ N, 89°45.162′ E, alt 2667 m, 10 Aug. 2016, leg. W. Klein, slide mounted (NBC). Paratypes: Same site and date, 2 ♂♂, 10 ♀♀ (2 ♂♂, 4 ♀♀ slide mounted, 6 ♀ in liquid) (NBC); Bhutan 02, E of Takchu, springs in meadow, 27°21.492′ N, 89°17.173′ E, 2907 m a.s.l., 07 Aug. 2016, leg. W. Klein, 1 ♂, 1 ♀ slide mounted, 2 ♀♀ in liquid; Bhutan 03, Dumcho, Haa river, pool 3, 27°21.883′ N, 89°17.738′ E, 2675 m a.s.l., 07 Aug. 2016, leg. W. Klein, 1 ♂ slide mounted.
Other material – Bhutan 03, Dumcho, Haa river, pool 3, 27°21.883′ N, 89°17.738′ E, 2675 m a.s.l., 07 Aug.2016, leg. W. Klein (Figure 6A), 1 dn slide mounted; Nepal 04, Surke Khola, stream, 27°40.327′ N, 86°43.297′ E, 2290 m a.s.l., 11 Apr. 2016 leg P.W. Veel (Figure 6D), 1 ♀ slide mounted (juvenile, grading to L. veeli n. sp. possibly due to incomplete sclerotization); Nepal 06, Sertu Khola stream, 27°33.899′ N, 86°35.752′ E, 2563 m a.s.l., 14 Apr. 2016 leg P.W. Veel (Figure 6B), 1 ♀ slide mounted; Nepal 09, Bhujekhung, Stream, 28°31.709′ N, 83°01.598′ E, 3100 m a.s.l., 10 Oct. 2016 leg P.W. Veel 1 ♂, 2 ♀♀ slide mounted; Nepal 10, Rupgad, Thuli Beri River, 28°58.460′ N, 82°51.991′ E, 2067 m a.s.l., 01 Nov. 2016 leg P.W. Veel, 2 ♀♀ slide mounted.
Diagnosis — Integument lineated, with lines in parts of the dorsum shortened and fusing to form a reticulate pattern; legs without swimming setae; P-3 basally relatively narrow (ratio maximum H/basal H 1.3-1.4), with 5 long setae, distal setae nearly equidistant or centrodistal seta closer to dorsodistal seta (A/B 1.3-2.1); P-4 maximum H in central part, insertions of setae dividing ventral margin in general into a short proximal, and subequal central and distal sector(s) (17-27/33-48/26-50 %).
Description — Integument lines surrounding longish fields of fine porosity, in parts of the dorsum shortened and fusing to form a characteristic reticulation, extension of this reticulation highly variable, in most specimens covering most of the dorsum, in others restricted to small dorsocentral fields. Cx-I medially distinctly longer than Cx-II (ratio 1.1-1.4), posterior margin W 25-35. Posterolateral margin of Cx-IV equally rounded or with a nearly straight sector on the level of glandularia V3. These and setae V1 not fused to coxal shield, but V3 often very close to Cx-IV margin (Figure 2D). Proportions and setation of distal segments of I- and IV-L as in Figure 2A-B; distal 2/3 of the ventral surface of IV-L-5 with more than 10 setae inserting close to each other, IV-L-6 slightly bowed, with 2-3 fine and short ventral setae; IV-L- claw L 55-67. Genital field (Figure 2E) with lateral margins of genital flaps strongly diverging from anterior to posterior end, medial setae (♂: < 30, ♀: ca 15), in general in a single row (posteriorly doubled in a few ♂♂). Excretory pore smooth. Gnathosoma ventral margin in distal part nearly straight. P-2 relatively robust (L/H 1.37-1.54), ventral seta inserted 7-10 from distoventral edge, basally strong, distinctly shorter than L of segment (Figure 2C); P-3 slender, in basal part narrower than distally, dorsal and ventral diverging distally, 2 of the 5 long setae arranged near the centre of the dorsum (close to, or clearly away from each other), centrodistal seta closer to dorsodistal one, distoventral seta near segment edge; P-4 with maximum H in centre or proximal part, mediodistal peg seta strong, pointed; proximoventral seta rather close to proximal segment edge, distoventral seta halfway between proximoventral seta and distal segment edge (in one exceptional case, the central sector is enlarged (26/48/26 %).
Etymology — Named after Rajan Khatri (Kathmandu), for his assistance with the collecting of the material.
Discussion — From the Asian fauna the only Lebertia (s.str.) species reported to have a reticulated integument is L. acuta Sokolow, 1934, described from the Ussuri catchment. This species differs from L. khatrii n. sp. in the position of the centrodistal of P-3 which is very close to the dorsodistal one (A/B 3.0). The same is true for L. hissarica Sokolow, 1948, a species from Tadjikistan that might have a similar integument structure (but not described explicitly, ratio A/B < 3.0). In the European fauna, two Lebertia s.str. species are known to have a reticulated integument, both differing from the new species, among others, in the position of ventral setae on P-4: Lebertia salebrosa Koenike, 1908, in comparison with the new species with a distinctly finer integument lineation locally producing a rather irregular reticulation, is generally minor in dimensions (e.g., P-4 L ♂ < 115, ♀ < 130), a farther distance between P-3 ventro- and centrodistal setae (ratio A/B 2.2-4.0), and the two proximal ventral sectors of P-4 shorter than the distal one (25/25/50 %), rarely equal in size (33/33/33 %). Specimens of L. reticulata Koenike, 1919 differ in presence of one swimming seta each on III-/IV-L-5, and a row of strong ventral setae on IV-L-6. Furthermore, their P-4 is club-shaped (maximum H in distal part) and both ventral setae insert in the distal segment sector.
Several specimens from Nepal do not show a distinct integument reticulation, but a pattern of longer or shorter lines similar to the situation in L. veeli n. sp., but they agree with the diagnosis in palp morphology.
Lebertia (Lebertia) disparilis n. sp.
ZOOBANK: DFB7EDBE-F312-4393-92A6-DFA3AE09BBEF ![]()
(Figures 3A-D, 6C)
Type series — Holotype ♂, Nepal 05, Garden stream, Nunthala (Figure 6C), 27°35.343′ N, 86°38.529′ E, 1955 m a.s.l., 13 Apr. 2016 leg P.W. Veel (RMNH). Paratype: ♂, Nepal 07, Bhujekhung, Stream, 28°31.714′ N, 83°01.648′ E, 3110 m a.s.l., 10 Oct. 2016 leg P.W. Veel, slide mounted (RMNH).
Diagnosis — (male, female unknown) Integument with a very fine reticulation; legs without swimming setae; palp with a stout, dorsally strongly curved P-2 (L/H 1.4), P-3 with 5 long setae, centrodistal seta closer to ventrodistal seta (A/B 0.2-0.6); P-4 long and slender (L/H 4.0-4.3, L ratio P-2/-4, 0.60-0.66), maximum H in proximal part, insertions of setae dividing ventral margin in three unequal sectors (the central the longest, the distal the shortest: 30-33/47-49/19-23 %).
Description — Male: Integument meshes rather fine (diameter ca. 3 µm), locally slightly elongated (dorsally arranged longitudinally, posteroventrally transverse. Cx-I and -II subequal in medial length (ratio 0.9-1.1), posterior margin W 25-30. Coxal shield extending to lateral idiosoma, subcircular in outline, also at posterior margin Cx-IV (Figure 3A); posteromedial edge of Cx-IV subrectangular (in the juvenile holotype ♂) or slightly projecting to embrace genital field (paratype), V3 and V1 not fused to coxal shield. Proportions and setation of distal segments of I- and IV-L as in Figure 3C-D; IV-L-6 slightly bowed, with 2-3 fine and short ventral setae (Figure 3D); IV-L claw L 47. Genital field compact with lateral margins of genital flaps nearly straight and diverging in anterior half, rounded in centre and posterior part, medial setae (about 20) in a single row, in addition, rather numerous (about 10) equidistant lateral setae. Excretory pore smooth. Gnathosoma ventral margin in proximal half and proximally to mouth opening nearly straight, in the distal 2/3 strongly bowed. P-2 relatively robust (L/H 1.41-1.43), dorsal margin strongly bowed, ventral seta inserted about 10 from distoventral edge, shorter than L of segment (Figure 3B); P-3 slender, slightly curved, dorsal and ventral diverging distally, 2 of the 5 long setae away from each other dorsomedially in proximal part, centrodistal seta closer to ventrodistal one, distoventral seta near segment edge; P-4 slender with maximum H in proximal part from here to tip dorsal and ventral margin equally converging, one dorsal hair-like seta near proximal segment edge, all others in a group near distal edge, mediodistal peg-like seta very fine, tip obtuse; proximoventral seta at about from ⅓ ventral margin, distoventral seta away, less than ¼ ventral margin from distal segment edge; P-5 short.
Etymology — Named for being morphologically very different from all other species of the genus.
Discussion — Lebertia disparilis n. sp. is extremely distinct, its palp has no similarity to any Lebertia species hitherto described. The palp of all known representatives of the subgenus Lebertia s.str. has a stouter P-4 (L/H < 4.0).
Subgenus Mixolebertia Thor, 1906
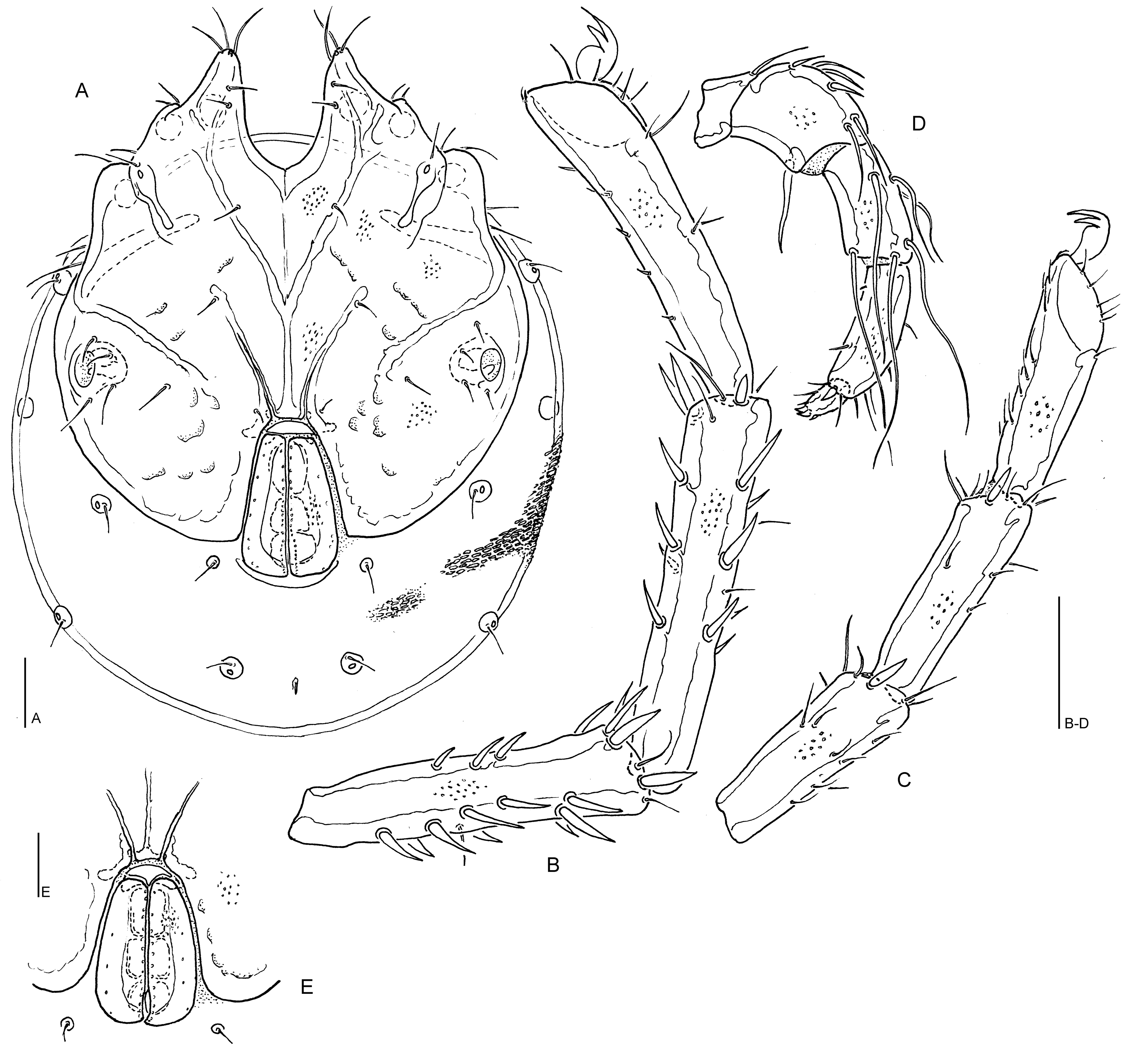
Lebertia (Mixolebertia) veeli n. sp.
ZOOBANK: 64E681D6-7F30-47EA-8A44-1B75481690A1 ![]()
(Figures 4A-D, 6D)
Type series — (all NBC) – Holotype ♂, Bhutan 01, Bhutan, Royal Botanical Park Lampelri, 27°30.450′ N, 89°45.162′ E, 2667 m a.s.l., 10 Aug. 2016, leg. W.Klein, slide mounted. Paratypes: Same site and date, 7 ♂♂, 4 ♀♀ (3 ♂♂, 3 ♀♀ slide mounted, 4 ♂♂, 1 ♀ in liquid).
Other material (all RMNH) – Nepal 04, Surke Khola, stream (Figure 6D), 27°40.327′ N, 86°43.297′ E, 2290 m a.s.l., 11 Apr. 2016, leg P.W. Veel, 1 ♀ slide mounted; Nepal 08, Laina Odar, Stream Tarap Khola, 28°55.616′ N, 83°02.627′ E, 3330 m a.s.l., 18 Oct. 2016, leg P.W. Veel, 1 ♀ slide mounted; Nepal 10, Rupgad, Thuli Beri River, 28°58.460′ N, 82°51.991′ E, 2067 m a.s.l., 01 Nov. 2016, leg P.W. Veel, 2 ♂ slide mounted.
Diagnosis — Integument lineated, lines short and thick; legs without swimming setae; P-3 basally thick (ratio maximum H/basal H 1.1-1.2), with 5-7 (in most cases, 6) long setae, centrodistal seta close to dorsodistal seta (A/B 2.2-4.7); P-4 maximum H in central part, insertions of setae dividing ventral margin in general into a very short proximal, and subequal central and distal sector(s) (15-20/40-44/38-43 %).
Description − Integument lines short and irregularly undulating, arrangement in centre of dorsum longitudinal, posterodorsally and ventrally transverse. Cx-I medially longer than Cx-II (ratio 1.1-1.5), posterior margin rather wide (40-50). Posterolateral margin of Cx-IV equally rounded or with a nearly straight sector on the level of glandularia V3. These and setae V1 not fused to coxal shield, but often V3 very close to Cx-IV margin (Figure 4A). Proportions and setation of distal segments of I- and IV-L as in Figure 4B-C; distal ⅔ of the ventral surface of IV-L-5 with less than 10 scattered setae, IV-L-6 slightly bowed, with 2-3 fine and short ventral setae (Figure 4B); IV-L claw L 70-80. Genital field with lateral margins of genital flaps strongly diverging from anterior to posterior end, medial setae (♂: < 30, ♀: ca 15) in a single row (Figure 4E). Excretory pore smooth. Gnathosoma ventral margin in distal part nearly straight. P-2 relatively robust (L/H 1.39-1.52), ventral seta inserted ca. 15 from distoventral edge, basally strong, shorter than L of segment (Figure 4D); P-3 slender (L/H 2.05-2.20, in basal part not narrowed, dorsal and ventral margins only very slightly diverging distally, 2 of the 6 (5-7) long setae arranged near the centre of the dorsum, 1–2 further ones halfway between base and centre, centrodistal seta near dorsodistal seta, distoventral seta set off from distoventral segment edge); P-4 L/H 2.9-3.1, with subparallel dorsal and ventral margins, mediodistal peg seta strong, pointed, proximoventral seta rather close to proximal segment edge, distoventral seta halfway between proximoventral seta and distal segment edge (in one exceptional case, the central sector is enlarged (18/58/24%).
Discussion — Among the Asian Mixolebertia species lacking swimming setae and with a smooth excretory pore, L. veeli n. sp. shares several important character states with two species described from Magadan (Siberia), L. mamolinensis Tuzovskij, 1982 and L. remota Tuzovskij, 2011 (''L. remotus'' Tuzovskij, 2011). Both have the medial suture Cx-I only slightly longer than suture Cx-II (mL ratio 1.0-1.5), a relatively stout palp (L/H P-3 < 2.3, P-4 < 3.2), and the central distomedial seta P-3 close to the dorsal one. Lebertia mamolinensis differs from L. veeli n. sp.in the arrangement of the setae on P-3 and P-4 (P-3: one of the medial setae shifted to the distal margin, resulting in presence of 4 mediodistal setae at this segment; P-4 ventral setae equidistant). Lebertia remotus is more similar, with agreement also in most aspects of proportions and setation of palp. A minor difference concerns the arrangement of P-4 ventral setae, with in L. remotus the proximoventral seta more away from the proximal segment edge, the distoventral one closer to distal segment edge, sectors 25-45-29% (but see above for the variability of this character in L. veeli n. sp.). The most important diagnostic character to separate L. veeli n. sp. from L. remota is the integument structure with short strong lines (in L. remota finely striated).
Subgenus Septlebertia Imamura, 1954
Lebertia (Septlebertia) altomontana n. sp.
ZOOBANK: F0BDD504-3FBA-41B1-8B9B-9A43B06147AD ![]()
(Figures 5A-D, 6E-F)

Type series — (all RMNH) – Holotype ♀, Nepal 01, stream S of Gokyo (Figure 6E), 27°56.860′ N, 86°41.853′ E, 4745 m a.s.l., 7 Apr. 2016 leg P.W. Veel, slide mounted (RMNH). Paratype ♀ Nepal 02, stream Dudh Kosi Nadi, Phortse (Figure 6F), 27°50.817′ N, 86°44.699′ E, 3596 m a.s.l., 9 Apr. 2016 leg P.W. Veel, slide mounted (RMNH).
Material compared – Lebertia convergella Tuzovskij, 1990, ♂, ♀ Russia, Magadan, snow valley stream, 15 Jun. 1982, leg Tuzovskij.
Lebertia imamurai Viets, 1956, ♂, ♀, Japan, Hokkaido, Kan-noko ike spring, 12 Aug. 2002, leg. Matsumoto; ♂, ♀, Hokkaido, Youtei-Hukiage spring, 14 May 2002, slide 100-2/3, leg. Matsumoto.
Diagnosis — Female (male unknown): Integument lineated; legs without swimming setae; palp with a dorsally strongly curved, ventrally weakly concave P-2 (L/H 1.5-1.6), P-3 with 6 long setae in three groups, each side by side: 2 dorsally, in proximal quarter, two dorsodistally and two ventrodistally; P-4 rather short (L/H 3.0-3.3, L ratio P-2/-4, 0.85-0.95), maximum H in proximal part, insertions of setae dividing ventral margin in sectors increasing ln L from base to tip: 21-27/29-34/43-45 %).
Description — Female: Integument densely covered by short, irregular, at both ends pointed lines (dorsally arranged longitudinally, posteroventrally transverse). Cx-I medially slightly to distinctly longer than Cx-II (ratio 1.2-1.8), posterior margin wide (about 50). Coxal shield in lateral part, on the level of leg insertions, equally rounded, posterolateral margin of Cx-IV in the area facing the unfused V3 nearly straight, towards the genital bay more rounded, posteromedial edges obtuse, V1 unfused (Figure 5B). Proportions and setation of distal segments of I- and IV-L as in Figure 5C-D; I-L-6 bowed, proximally slender, ventral and dorsal margins strongly diverging to form an elevated area of claw insertion, with 2-3 fine and short ventral setae; IV-L claw large (L 75-85). Genital field with lateral margins of genital flaps very slightly convex, continuously diverging from anterior edge towards the rounded posterior margin, each flap with about 25 medial, and 5 lateral setae. Excretory pore smooth. Gnathosoma ventral margin in proximal third nearly straight, from here to distal end equally rounded, a fine kink proximal to mouth opening. Palp with a dorsally strongly curved, ventrally weakly concave P-2 (L/H 1.5-1.6), ventral seta basally thick, shorter than L of segment, insertion associated with a blunt projection about 20 from distoventral edge (Figure 5A); P-3 nearly parallelogram-shaped, dorsal and ventral margins diverging distally, the two proximodorsal setae close to segment base, distance between the two groups of distal setae about two times the distance between the dorsodistal resp. ventrodistal setae; P-4 stout and short with maximum H in proximal part, from here to tip dorsal and ventral margin slightly converging; two dorsal hair-like seta in area of segment centre, the others in a group near distal edge, mediodistal peg-like seta robust, pointed; P-5 not shortened.
Etymology — Named for its occurrence high in the mountains.



Discussion — Lebertia altomontana n. sp. differs from most known Lebertia species in the setation of palp segment P-3 (with six medial setae arranged in three groups – proximodorsally, dorsodistally, and ventrodistally, no setae shifted to lateral surface), combined with a P-4 with ''normally'' arranged ventral setae (distanced from each other, not side-by-side). A similar P-3 setation is reported for L. gibberipalpis Sokolow, 1948 from Tadjikistan, L. noshaqensis Imamura, 1966 from Afghanistan, and L. xuanensis Wang et al., 2020 from Central China. Lebertia gibberipalpis differs from L. altomontana n. sp. in the lower mL ratio Cx-I/II (1.1), a distinctly stouter palp (L/H P-2, 1.27; P-3, 1.71; P-4, 2.56), and both ventral setae on P-4 shifted to the proximal half of the segment (ventral sectors 1:1:2). Lebertia noshaqensis requires a redescription. From the original description we can deduce that it differs from L. altomontana n. sp. in minor dimensions (e.g., genital flap L < 200, P-4 L < 130), a subrectangular P-3 proximally only slightly narrower than distally, and the position of the distoventral seta on P-4 (distance from distoventral segment edge < segment H). Lebertia xuanensis differs from L. altomontana n. sp. in the dorsal setae on P-3 more away from each other, ventral setae on P-4 very close to each other (sectors 2:1:2), and a by far lower number of ventral setae on IV-L-4/-5. It is furthermore characteristic in rather tiny dimensions (e.g., coxal field L 455, P-2-3-4 L 76-76-82). This observation remains vaild if a discrepancy between measurements and figures in the original description is taken in account – from the bar in Figure 2C, higher, but in comparison with other species still quite low, values are deduced for palp segments (P-2-3-4 L 90-100-110). Also L. mamolinensis Tuzovskij, 1982 from Magadan (Siberia) has 4 of the six P-3 setae at the distal segment margin, but the setae are not as clearly grouped (distoventral seta isolated from the other three distal setae). Lebertia mamolinensis differs furthermore in a lower Cx-I/II mL ratio (1.0-1.2), stouter P-3 (L/H < 2.1), and equidistant ventral setae on P-4.
Attributing the new species to Septlebertia Imamura, 1954 might surprise as it differs from the diagnosis of the subgenus just in the name-giving setae number on P-3 (six, not seven). However, in the type species L. (S.) imamurai K. Viets, 1956, four mediodistal setae are likewise arranged in a dorsal and a ventral pair. Furthermore, revision of new material evidences a wide variability of P-3 setation also in L. imamurai: A specimen with a typical palp on one side had nine medial setae (4 dorsal, two dorsodistal, and 3 ventrodistal) on the facing P-3. In the slender shape of palp segments, the new species agrees with L. (S.) dmitrii Semenchenko & Tuzovskij, 2010. Both species differ in this regard from L. imamurai and L. convergella Tuzovskij, 1990, with L/H P-2 < 1.5, P-3 < 2.0, P-4 < 3.0 . In palp proportions, e.g., slender segments P-2 (L/H < 1.5) and P-3 (L/H < 2.0), the new species is similar to L. boldyrbekensis Tuzovskij, 2021 from Kazachstan from which it differs in the lower number of P-3 setae and a less curved IV-L-6.
Eolebertia Gerecke, 2009, the other Lebertia subgenus with doubled distoventral setae, differs from Septlebertia species in the position of the distal setae (at least one of them shifted to the lateral surface). Furthermore, Eolebertia differs from all other subgenera in the side-by-side arrangement of the ventral setae on P-4. As far as we know, this subgenus is not represented in the Asian fauna. Lebertia exceedia Wang, Wang, Jin & Guo, 2020, a species described from China, has been erroneously attributed to Eolebertia. Having a total number of 8 setae on P-3 (all distal setae on this segment on the medial surface), and ventral setae of P-4 away from each other, at present state of taxonomy L. exceedia is a typical representative of Septlebertia.
Acknowledgements
The authors are indebted to Piet Veel (Heemstede), Rajan Khatri (Kathmandu), Wim Klein and Oscar Vorst (RMNH) for collecting of the material in Nepal and Bhutan, respectively, and to Petr Tuzovskij (Borok) and Noriko Matusomto (Sapporo) for the loan of material. Two anonymous reviewers helped to improve this paper with careful and constructive criticism.
References
- Cook D.R. 1967. Water mites from India. Mem. Amer. Entom. Inst., 9: 1-411.
- Gerecke R. 2009. Revisional studies on the European species of the water mite genus Lebertia Neuman, 1880 (Acari: Hydrachnidia: Lebertiidae). Abh. Senck. Ges. Naturfsch., 566: 1-144.
- Kumar N., Kumar K., Pešić V. 2007a. Two new species of Sperchon Kramer (Acari: Hydrachnidia: Sperchontidae) from the Garhwal Himalayas (India). Syst. Appl. Acarol., 12: 31-36. https://doi.org/10.11158/saa.12.1.5
- Kumar N., Kumar K., Kumar S., Pešić V. 2007b. Monatractides tuzovskyi sp. nov. (Acari: Torrenticolidae), a new water mite from the Garhwal Himalayas (India). Acarina, 14: 181-183.
- Lundblad O. 1934. Report on Hydracarina. Mem. Connecticut Acad. Arts, Sci., 10: 85-118.
- Panesar A.R. 2004. Evolution in water mites (Hydrachnellae, Actinedida, Acari). Bonner zool. Monogr., 52: 1-144.
- Panesar A.R., Gerecke R. 1994. A new Partnunia species (Acari, Actinedida, Hydryphantidae) from Keylong (Himalaya, India). Aquatic Insects, 16: 241-247. https://doi.org/10.1080/01650429409361560
- Pešić V., Chatterjee T., Kumar Das M., Bordoloi S. 2012. Two rare water mit species (Acari: Hydrachnidia) from the streams of the Indian eastern Himalayan region. Syst. Appl. Acarol., 17: 458-464. https://doi.org/10.11158/saa.17.4.15
- Pešić V., Gerecke R. 2008. A new water mite (Acari: Hydrachnidia, Sperchontidae) from the Himalaya Mountains (Northern India). Vestn. zool., 42: 9-12. https://doi.org/10.2478/v10058-008-0006-2
- Pešić V., Kumar N., Kumar K. 2007a. Two new species of water mites of the family Hygrobatidae (Acari: Hydrachnidia) from the Garhwal Himalayas (India). Syst. Appl. Acarol., 12: 161-166. https://doi.org/10.11158/saa.12.2.11
- Pešić V., Kumar N., Kumar K. 2007b. A new species of Monatractides (Acari: Hydrachnidia: Torrenticolidae) and new records of other torrenticolid water mites from the Garhwal Himalayas (India). Syst. Appl. Acarol., 12: 205-212. https://doi.org/10.11158/saa.12.3.5
- Pešić V., Panesar A. 2008. Stdies on water mites (Acari, Hydrachnidia) from the Himalyas, I. The water mite genus Feltria Koenike, with description of eight new species. Zootaxa, 1758: 1-28. https://doi.org/10.11646/zootaxa.1758.1.1
- Pešić V., Smit H. 2007. First record of water mites (Acari: Hydrachnidia) from Bhutan, with description of two new species. Zootaxa, 1613: 45-56. https://doi.org/10.11646/zootaxa.1613.1.3
- Pešić V., Smit H., Negi S., Bahaguna P., Dobriyal A.K. 2020a. Torrenticolid water mites of India with descriptions of three new species (Acari: Hydrachnidia, Torrenticolidae). Syst. Appl. Acarol., 25: 255-267. https://doi.org/10.11158/saa.25.2.7
- Pešić V., Smit H., Bahaguna P. 2020b. A new species of Kongsbergia from the Western Himalaya with a key to the species of the genus of India (Acari: Hydrachnidia). Ecol. Monten., 27: 35-38. https://doi.org/10.37828/em.2020.27.4
- Pešić V., Smit H., Bahaguna P. 2019a. New records of water mites (Acari: Hydrachnidia) from the Western Himalaya with the description of four new species. Syst. Appl. Acarol., 24: 59-80. https://doi.org/10.11158/saa.24.1.5
- Pešić V., Smit H., Bahaguna P. 2019b. New records of water mites (Acari: Hydrachnidia) from the Western Himalaya and description of three new species from Asia. Syst. Appl. Acarol., 24: 1868-1880. https://doi.org/10.11158/saa.24.10.6
- Walter C. 1928. Zur Kenntnis der Mikrofauna von British Indien. Rec. Indian Mus., 30: 57-108.
- Wang J., Jin D-C., Yi T., Guo J. 2016 Water mites of the subgenus Lebertia (Lebertia) Neuman from China (Hydrachnidia: Lebertiidae). Zool. Syst., 41: 54-63.



2022-01-13
Date accepted:
2022-03-07
Date published:
2022-03-11
Edited by:
Mąkol, Joanna

This work is licensed under a Creative Commons Attribution 4.0 International License
2022 Gerecke, Reinhard and Smit, Harry
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)