



Communities of predatory mites (Phytoseiidae and Stigmaeidae) in different environments of the Brazilian Pampa
Toldi, Maicon1 ; de Freitas, Elisete Maria2 ; da Silva, Vinícius Leão3 ; Caumo, Monique4 ; Ferla, Júlia Jantsch5 ; Orlandi, Carla Roberta6 ; Da-Costa, Tairis7 ; Johann, Liana8 and Ferla, Noeli Juarez9
1Universidade do Vale do Taquari - Univates, Lajeado, CEP 95913-528, Rio Grande do Sul, Brasil.
2Universidade do Vale do Taquari - Univates, Lajeado, CEP 95913-528, Rio Grande do Sul, Brasil.
3Universidade do Vale do Taquari - Univates, Lajeado, CEP 95913-528, Rio Grande do Sul, Brasil.
4Universidade Federal do Rio Grande do Sul, Porto Alegre, CEP 90040-060, Rio Grande do Sul, Brasil.
5Universidade Federal de Viçosa, Viçosa, CEP 36570-900, Minas Gerais, Brasil.
6Universidade do Vale do Taquari - Univates, Lajeado, CEP 95913-528, Rio Grande do Sul, Brasil.
7✉ Universidade Federal do Rio Grande do Sul, Porto Alegre, CEP 90040-060, Rio Grande do Sul, Brasil.
8Universidade do Vale do Taquari - Univates, Lajeado, CEP 95913-528, Rio Grande do Sul, Brasil.
9Universidade do Vale do Taquari - Univates, Lajeado, CEP 95913-528, Rio Grande do Sul, Brasil & CNPq Researcher.
2021 - Volume: 61 Issue: 1 pages: 20-30
https://doi.org/10.24349/acarologia/20214413Original research
Keywords
Abstract
Introduction
The Pampa environment is present in several countries (Argentina, Brazil, Paraguay and Uruguay) and is characterized by vegetation composed mainly of grasses and herbaceous species (Carvalho et al. 2009). South Brazilian native pastures occupy about 13.7 million hectares and support very high levels of biodiversity, it provides several environmental services and forage potential for livestock (Pillar et al. 2009). In Brazil, the Pampa vegetation comprises a mosaic riparian forests within a dominant grassland habitat (Behling et al. 2005). This habitat is under considerable human pressure, with less than 40% of its original native vegetation remaining (MMA 2010).
The high diversity of fauna and flora in the Pampa provides environmental services, conserves water resources, pollinators, genetic resources, and foraging for livestock (Pillar et al. 2009). Some human activities favor the maintenance of the grassland environment, e.g. grazing activities and fire that prevent the expansion of forest formations over grasslands (Behling et al. 2007). However, grasslands have been recently replaced by grain crops and Eucalyptus saligna Smith (Myrtaceae) plantations (Pillar et al. 2009). These modifications in land use can have deleterious effects on the diversity and distribution of organisms in this environment, such as abundance and diversity of collembolans and edaphic mites (Rieff et al. 2016).
Several factors might affect environmental recovery and analyses are required to compare different mature forest areas and assess changes in species composition and regeneration of degraded areas (Coote et al. 2013; Derroire et al. 2016). Since it is not possible to totally analyze the environment with high biological diversity such as the Pampa, the use of ecological indicators is an alternative way for the diagnosis of environmental quality (Coote et al. 2013). Studies indicate that soil mites might be environmental indicators of both native areas and areas of exotic monocultures (Rieff et al. 2016). These ecological indicators are used in decision-making processes regarding management and public policies for environmental recovery and conservation (Niemi and McDonald 2004).
Plant mites represent one of the most diverse and abundant groups in terrestrial ecosystems (Walter and Proctor 2013). In spite of that, studies that analyze their potential as indicators of both anthropised environments and preserved environments are scarce (Rocha et al. 2015). One of the most abundant groups are the predatory mites of the families Phytoseiidae and Stigmaeidae, which are abundant on plants worldwide (Gerson et al. 2003) and are often employed for biological control issues (i.e. Tixier 2018). The number of Phytoseiidae species is high in the Neotropical region (McMurtry et al. 2013; Araújo and Daud 2018; Araújo and Kollár 2019), with 2,798 species reported in the world (Demite et al. 2020). Stigmaeidae live on plants and soil, and this family comprises 577 species described worldwide (Fan et al. 2016). In the state of Rio Grande do Sul, stigmaeids live in several agroecosystems and are considered the second most abundant predatory family after Phytoseiidae (Ferla and Moraes 2002). Several studies provide information on the occurrence of predatory mites on plants, but little is known about the parameters that explain this occurrence (i.e. Tixier 2018).
The aim of this study was to evaluate the communities of predatory mites (Phytoseiidae and Stigmaeidae) in grasslands, modified grasslands, and riparian forests in the Pampa of the South of Brazil. The main hypothesis, herein tested, is that forests have different communities of predatory mites compared to grassland environments, in a way that the forest area presents an assembly of specific predatory mites and the diversity within grassland areas is lower.
Material and methods
Study area
In the municipality of Pântano Grande, Brazil (30°19'36.59'' S, 52°29'24.83'' W, 100 meters above sea level), the sampling units (sites) are located in permanent preservation areas composed of native species. Around these sites, there are plantations of Eucalyptus sp. These areas belong to the ''Shrub Field vegetation type'', located in the Central Depression of the Pampa. This area is classified as Cfa, i.e., with a well-delimitated summer and winter seasons, and annual rainfall varying from 1,600 to 1,900 mm (Peel et al. 2007). The soil is Red dystrophic Argisol (Streck et al. 2018). The areas herein investigated were used for agriculture and cattle breeding until 2006, when Eucalytus sp. plantations started and environmental recovery areas were defined.
The samplings were carried out in nine sites included in three environments, with areas larger than 0.5 ha and for each type of environment. The three types of environments were (i) Riparian Forest, with tree vegetation occurring in the margins of water resources. Riparian forests composed of a vegetation of Seasonal Semi-deciduous Forest (high litter-fall in autumn); (ii) Grazing Exclusions, with environmental recovery measures, fenced for six years, with patches of shrub vegetation; and (iii) Grassland, with compacted soil and herbaceous vegetation, with no fencing or recovery measures, and access for cattle to grazing.
Three surveys were carried out (September 2017, November 2017 and February 2018), starting 20 meters away from the edge of the area to avoid border effects (Rodrigues et al. 2010). Twelve native plant species were selected at each site based on studies of plant communities in the same areas (Caumo 2018). In total, 55 species of native plants were sampled, some plant species being sampled in different sites. The plant species selected for the present study were those that had the highest Importance Value Indices at each sampling unit, calculated using the mean of the sum of Relative Cover and Relative Frequency values. These data were provided by the Department of Horticulture and Forestry of the Agronomy School of Federal University of Rio Grande do Sul, Porto Alegre, RS and by the Laboratory of Botany of Univates, Lajeado, RS.
At each of the nine sites, 12 plant species were sampled with three replications over time, totaling 324 plant samples. Plant leaves were collected and kept in plastic bags within Styrofoam boxes (approximate temperature of 5 °C) until they were brought back to the laboratory. For each sample, mites were searched on leaves and stems for one hour, using a stereomicroscope Leica EZ4 HD, for standardizing the observation effort per plant. All mites retrieved were mounted on microscope slides in Hoyer's medium and kept at 40 °C for a period of seven days for clearing (Moraes and Flechtmann, 2008). Female specimens were identified at species level using a phase-contrast optical microscopy (Zeiss Axio Scope A1), based on identification keys (Moraes et al. 2004; Lindquist et al. 2009; Walter et al. 2009; Fan et al. 2016).
The plants were identified by the Department of Horticulture and Forests of the School of Agronomy of the Federal University of Rio Grande do Sul (UFRGS) and by the Botany Laboratory of the University of Vale do Taquari (Univates), with consultation specialists and herbariums.
Data analyses
Interactions between mites and plants were analyzed using the ecological network analysis using the ''network-level'' function of the Bipartite package of the R software (R Core Team, 2019). We evaluated connectivity and robustness, which expresses the proportion of all possible interactions between species of mites and plants and assess the resistance of the network to disturbances, respectively (Dunne et al. 2002; Blüthgen et al. 2006).
Predatory mite communities from the three environments were analyzed using a non-metric multidimensional scaling (NMDS), to assess dissimilarity in the assemblages of predatory mites in the different environments analyzed (Riparian Forest, Grazing Exclusions and Grassland). We also applied the Bray-Curtis dissimilarity index, through an abundance matrix. To detect significant differences in predatory mite communities between environments, we performed a similarity analysis (ANOSIM) using the software PRIMER-E, version 5.2.9 (Clarke and Gorley 2002).
Potential ecological indicator species were identified with the Indicator Value method (IndVal) (Dufrêne and Legendre 1997). The IndVal method is based on exclusivity for a habitat and frequency of occurrence within the same habitat. Calculations of Indval were performed for each species and site with the package `labdsv' v1.8-0 in R (R Core Team, 2019). The statistical significance of indicator values was measured using random iterative methods by relocating 10,000 samples within sample groups. Species were considered as indicators if their maximum value of Indval was 25% (Dufrêne and Legendre 1997).
Results
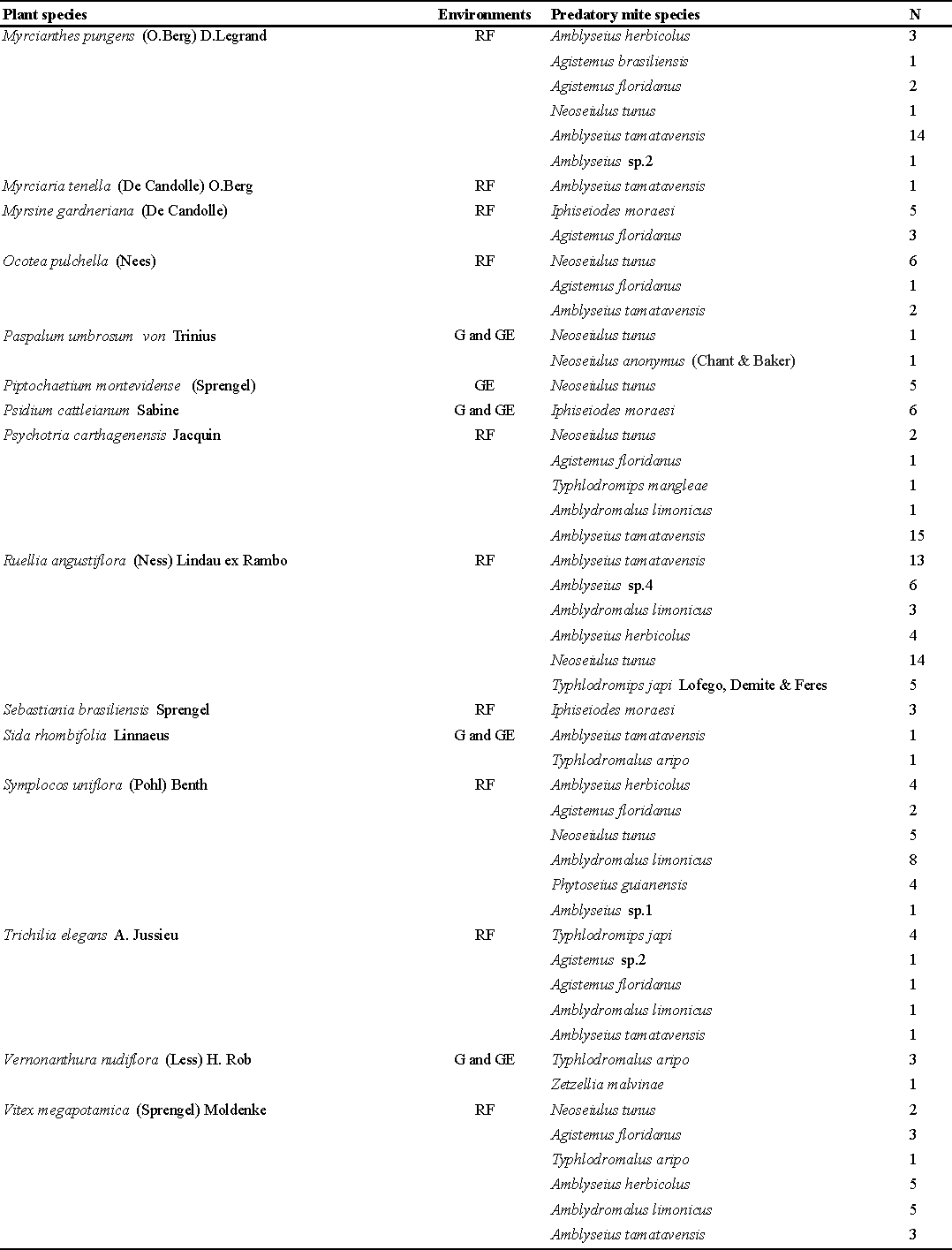
On the 55 plant species analyzed, predatory mites were observed on only 33 species in the three environments (Table 1). A total of 332 mite specimens were found, most of them belonging to the family Phytoseiidae (22 species, 293 specimens) followed by stigmaeids (6 species, 39 specimens). Riparian Forest was the environment with the highest number of specimens (243), followed by Grazing Exclusions (64) and Grassland (25). The most abundant species were represented by only five species: Neoseiulus tunus (De Leon) (64 specimens), Amblyseius tamatavensis Blommers (56 specimens), Amblyseius herbicolus (Chant) (48 specimens), Amblydromalus limonicus (Garman and McGregor) (30 specimens) and Agistemus floridanus Gonzalez-Rodriguez (25 specimens (Table 1).


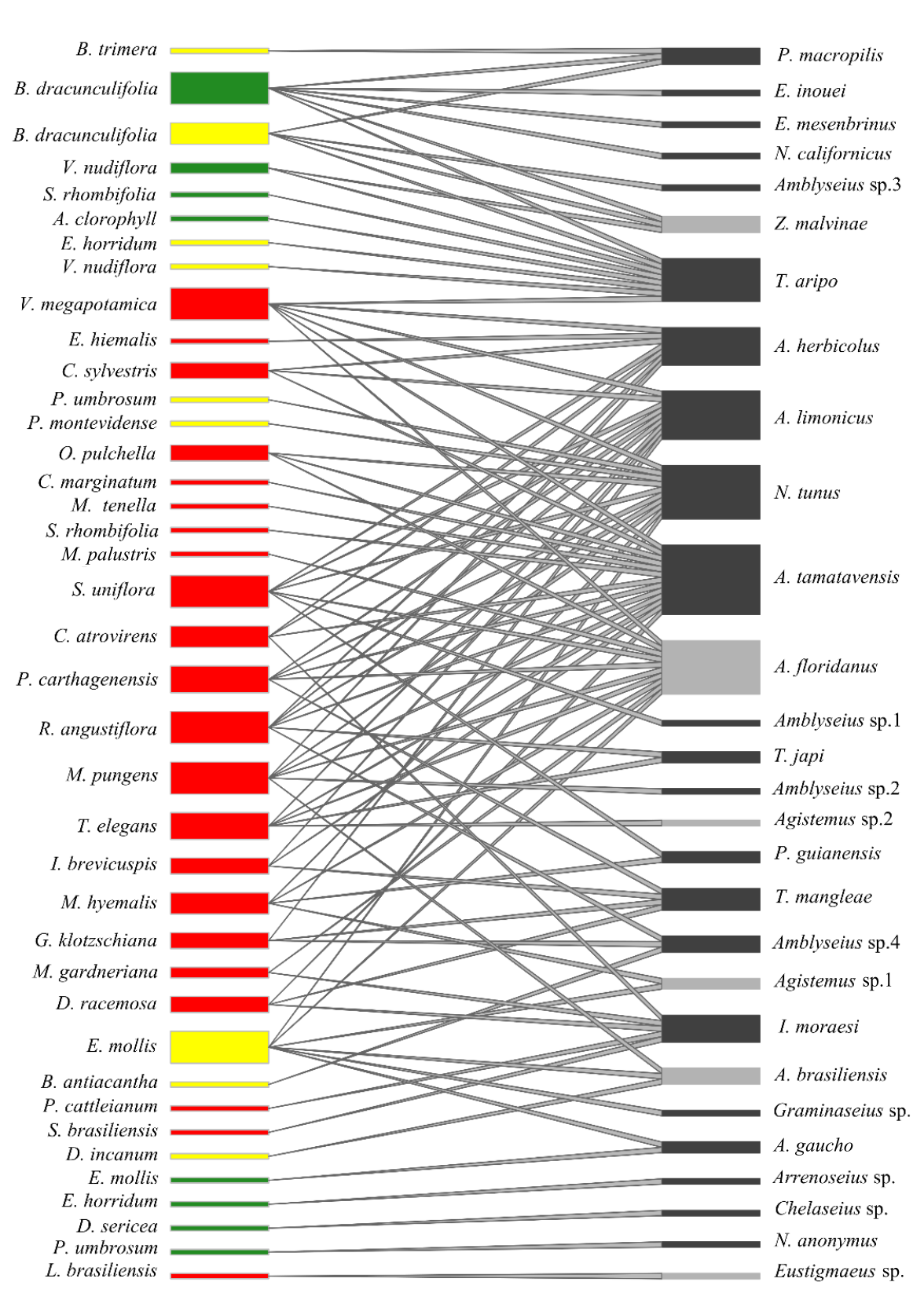
Networks of mites and plants showed 98 interactions (Robustness = 0.583; connectivity = 0.090) (Figure 1). Amblyseius tamatavensis and A. floridanus were the species with the highest number of interactions within the families Phytoseiidae and Stigmaeidae, respectively. Baccharis dracunculifolia De Candolle was the plant species showing the highest number of interactions (with six species in the Grassland environment: Euseius inouei Ehara and Moraes, Euseius mesembrinus (Dean), Neoseiulus californicus (McGregor), Phytoseiulus macropilis (Banks), Typhlodromalus aripo (De Leon) and Zetzellia malvinae Matioli, Ueckermann and Oliveira). Elephantopus mollis Kunth was the plant species with the highest number of interactions in Grazing Exclusion environment (with six species). Four plant species had high number of interactions in the Riparian Forest environment, Vitex megapotamica (Sprengel) (with six species), Symplocos uniflora (Pohl) (with six species), Ruellia angustiflora (Ness) (with six species), and Myrcianthes pungens (Berg) Legrand (with six species).
NMDS analysis ordinated the nine sites of the three environments, grouping together the Grassland and Grazing Exclusions, the forest remaining separated (Figure 2). The disturbance value provided by the scaling (stress value = 0.06) indicates that the ordination well fits the graphical representation. This result was confirmed by the ANOSIM analysis (global R = 0.462, P < 0.001), indicating that the three areas are characterized by different mite communities. Comparisons between Grassland and Grazing Exclusions did not show any differences (R = 0.092, P < 0.05), where difference was observed between Grassland and Riparian Forest environments (R = 0.45, P < 0.001), and between Grazing Exclusions and Riparian Forest environments (R = 0.46, P < 0.001). The Riparian Forest was the environment with the greatest predatory mite richness and abundance, and the Grassland and Grazing Exclusions areas presented more similar diversity and lower diversity and abundance of predatory mites.

Approximately 50% of mite species were detected as environmental indicators. Riparian Forest was the environment with the highest number of environmental indicator species, with A. tamatavensis, A. herbicolus, A. floridanus and A. limonicus, with IndVal higher than 0.50. Zetzellia malvinae and Arrenoseius gaucho Ferla, Silva and Moraes were detected as indicator species in Grassland environment and only one species, T. aripo, was detected as an indicator species in the Grazing Exclusions environment (Table 2).

Discussion
The results herein obtained support our initial hypothesis: mite communities are different between Riparian Forest and Grassland areas. Mite communities in Grasslands and modified grasslands areas were similar to each other's and diversity was lower than in the Riparian Forest area. Those results show that the Brazilian Pampa, because of diverse vegetation biome, shelter a high diversity of predatory mites associated with native plants, demonstrating the importance of this ecosystem as a reservoir of natural enemies.
Studies on mite diversity in the Brazilian Pampa are scarce, here we found 28 species of predatory mites (22 species of Phytoseiidae and 6 species of Stigmaeidae) on 33 plant species. In Brazilian Cerrado, Demite et al. (2017) found 34 phytoseiid species on 37 plant species. In Pantanal, Mendonça et al. (2019) found 35 phytoseiid species on 40 native plants and Rodrigues et al. (2020) found 11 phytoseiid species associated with Myrtaceae plants in the Caatinga. In the Brazilian Atlantic Forest, Ferla and Moraes (2002) found 23 phytoseiid species on 45 plant species; Castro and Moraes (2010) found 54 phytoseiid species on 187 species of plants; Demite et al. (2013) found 31 Phytoseiid species on two native plants in fragments of semi-deciduous seasonal forest; Rocha et al. (2015) reported 29 Phytoseiid species on 62 plants; Lofego et al. (2017) found 29 species on Cecropia pachystachya Trec. and Nuvoloni et al. (2015) found 29 Phytoseiid species on Hevea spp. from the Amazon Forest.
The richness of Phytoseiidae found in this study is very similar to that found in the Brazilian Caatinga and Cerrado and is lower than that observed in the Atlantic Forest and Amazon Forest. The richness of Stigmaeidae found in this study was higher than that observed in other studies (Ferla and Moraes 2002; Demite et al. 2013; Demite et al. 2015; Lofego et al. 2017; Noronha et al. 2020; Rodrigues et al. 2020), but similar to that found by Johann et al. (2014) in vineyard agro-ecosystems. Many plants sheltered more than one mite species and the interaction value was high with 98 interactions of 28 mite species. The robustness of the networks was similar to that found by Araujo and Kollár (2019). This can be explained by the fact that these mites are predators, as networks with phytophagous mites have lower connectivity (Araújo and Daud 2018). The assemblages of predatory mites of Riparian Forest contrasts with grassland and grazing exclusions. Thus, predatory mites can also be considered a meaningful indicator of environmental characteristics in the Brazilian Pampa. Amblyseius tamatavensis, A. herbicolus and A. floridanus are associated to Riparian Forest. Amblyseius tamatavensis and A. herbicolus are worldwide distributed (Demite et al. 2019) and A. floridanus only occurs in the American continent (Johann et al. 2013). Arrenoseius gaucho, collected to Grassland, has been described in this environment (Ferla et al. 2010), but there is no ecological information in literature regarding this species. Similar to Z. malvinae, which is related to Grassland, it is distributed throughout Brazil and occurs in rubber trees, vineyards, citrus, and coffee crops (Matioli et al. 2002; De Vis et al. 2006; Mineiro et al. 2006; Diehl et al. 2012). Typhlodromalus aripo was more associated with Grazing and Grazing Exclusions, this species was also associated to unstable environments with extreme windy conditions (Rocha et al. 2015).
Plant species that showed the highest number of predatory species could be selected in future studies on environmental indicators, especially to determine what are the factors affecting mite species occurrence. These plants occur throughout several countries and biomes (Ezcurra 1989; Patrício and Cervi 2005; Borges and Forzza 2008; Carvalho 2008; Cruz et al. 2016) and may be favorable to mites due to particular plant traits, such as trichomes and domatia, providing shelter and food (i.e. Tixier 2018). The main questions would be: the mite species occurrence is leaded by factors associated to environmental and /or plant features?
Predatory mites are important biological control agents of other mite groups and several species are used in many biological control programs (Gerson et al. 2003; McMurtry et al. 2013). Here, we found economically important biocontrol agents (MAPA 2018) particularly P. macropilis (9 specimens) and N. californicus (1 specimen). Other predators found in this study have also economic importance in biological control as shown in several studies: Agistemus floridanus (Ferla and Moraes 2003), A. limonicus (Vangansbeke et al. 2014), A. tamatavensis (Cavalcante et al. 2017), E. mesembrinus (Landeros et al. 2004) and T. aripo (Onzo et al. 2012).
Finally, we conclude that the Brazilian Pampa has many species of predatory mites, whose diversity is affected by the vegetation. The different environments of the Brazilian Pampa show a rich diversity of predators and can contribute to biological control programs. Thus, the monitoring of these areas is important to assess how these populations are maintained and evolve through time, especially in the ecological succession processes.
Acknowledgments
The authors are grateful to Celulose Riograndense and UNIVATES for their financial support. This study was partly financed by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Finance Code 001. The authors are grateful to the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) [productivity scholarship nº. 310035-2017-1. awarded to NJF].
References
Araújo W.S., Kollár J. 2019. First characterization of a highly specialized ecological network composed by gall-inducing mites and their host plants. Int. J. Acarol., 45: 223-226. doi:10.1080/01647954.2019.1591505
Araújo W.S., Daud R.D. 2018. Investigating effects of host-plant diversity on Brazilian mite richness in natural ecosystems. Syst. Appl. Acarol., 23: 1598-1613. doi:10.11158/saa.23.8.10
Behling H., Pillar V.D., Müller S.C., Overbeck G.E. 2007. Late Holocene fire history in a forest grassland mosaic in southern Brazil: Implications for conservation. Appl. Veg. Sci., 10: 81-90. doi:10.1111/j.1654-109X.2007.tb00506.x
Behling H., Pillar V.D., Bauermann S.G. 2005. Late Quaternary grassland (Campos), gallery forest, fire and climate dynamics, studied by pollen, charcoal and multivariate analysis of the São Francisco de Assis core in western Rio Grande do Sul (southern Brazil). Rev. Palaeobot. Palynol., 133: 235-248. doi:10.1016/j.revpalbo.2004.10.004
Blüthgen N., Menzel F., Blüthgen N. 2006. Measuring specialization in species interaction networks. BMC Ecol., 6: 1-12. doi:10.1186/1472-6785-6-9
Borges R.A.X., Forzza R.C. 2008. A tribo Astereae (Asteraceae) no Parque Estadual do Ibitipoca, Minas Gerais, Brasil. Boll. Bot. Univ. São Paulo, 26: 131-154. doi:10.11606/issn.2316-9052.v26i2p131-154
Carvalho P.E.R. 2008. Maria-Mole-do-Banhado (Symplocos uniflora). Embrapa Florestas Circular Técnica. p. 148.
Carvalho P.C.F., Batello C. 2009. Acesso à terra, produção pecuária e conservação de ecossistemas no bioma Campos: o dilema dos pastagens naturais. Pecuária Ciência, 1: 158-162.
Castro T.M.M.G., Moraes G.J. 2010. Diversity of phytoseiid mites (Acari: Mesostigmata: Phytoseiidae) in the Atlantic Forest of São Paulo. Syst. Biodiv., 8: 301-307. doi:10.1080/14772001003801375
Caumo M. 2018. Estudo de comunidades vegetais em áreas de preservação permanente associadas à silvicultura e à pecuária no bioma Pampa, Rio Grande do Sul [Master's Thesis]. Universidade Federal do Rio Grande do Sul, Porto Alegre, Rio Grande do Sul, Brasil.
Cavalcante A.C.C., Mandro M.E., Paes E.R., Moraes G. J. 2017. Amblyseius tamatavensis Blommers (Acari: Phytoseiidae) a candidate for biological control of Bemisia tabaci (Gennadius) biotype B (Hemiptera: Aleyrodidae) in Brazil. Int. J. Acarol., 43: 10-15. doi:10.1080/01647954.2016.1225816
Clarke K.R., Gorley R.N. 2002. Primer 5.2.9. [Internet]. Available in: https://www.primer-e.com/

Coote L., Dietzsch A.C., Wilson M.W., Graham C.T., Fuller L., Walsh A. T., Irwin S., Kelly D.L, Mitchell F.J.G., Kelly T.C., O$'$Halloran J. 2013. Testing indicators of biodiversity for plantation forests. Ecol. Indicators, 32: 107-115. doi:10.1016/j.ecolind.2013.03.020
Cruz A.P.O., Viana P.L., Santos J.U. 2016. Flora das cangas da Serra dos Carajás, Pará, Brasil: Asteraceae. Rodriguésia, 67: 1211-1242. doi:10.1590/2175-7860201667520
Demite P.R., Lofego A.C., Feres R.J. 2013. Mite (Acari; Arachnida) diversity of two native plants in fragments of a semideciduous seasonal forest in Brazil. Syst. Biodiv., 11: 141-148. doi:10.1080/14772000.2013.806368
Demite P.R., Feres R.J.F., Lofego A.C. 2015. Influence of agricultural environment on the plant mite community in forest fragments. Braz. J. Biol., 75: 396-404. doi:10.1590/1519-6984.14913
Demite P.R., Dias M.A., Cavalcante A.C.C., Ramos M.V.V., Lofego, A.C. 2017. Phytoseiid mites (Acari: Mesostigmata: Phytoseiidae) associated with Cerrado biome plants in Brazil, with description of a new species. Syst. Appl. Acarol., 22: 2141-2177. doi:10.11158/saa.22.12.9
Demite P.R., Moraes G.J., McMurtry J.A., Denmark H.A., Castilho, R.C. 2020. Phytoseiidae Database. [Internet]. Available online: www.lea.esalq.usp.br/phytoseiidae
Derroire G., Balvanera P., Castellanos-Castro C., Decocq G., Kennard D.K., Lebrija Trejos E., Leiva J.A., Odén P.C., Powers J.S., Rico-Gray V., Tigabu M., Healey J.R. 2016. Resilience of tropical dry forests-a meta-analysis of changes in species diversity and composition during secondary succession. Oikos, 125: 1386-1397. doi:10.1111/oik.03229
De Vis R.M.J., Moraes G.J., Bellini, M.R. 2006. Effect of air humidity on the egg viability of predatory mites (Acari: Phytoseiidae, Stigmaeidae) common on rubber trees in Brazil. Exp. Appl. Acarol., 38: 25-32. doi:10.1007/s10493-005-5444-8
Diehl M., Ferla N.J., Johann, L. 2012. Plantas associadas a videiras: uma estratégia para o controle biológico no Rio Grande do Sul. Arq. Inst. Biol., 79: 579-586. doi:10.1590/S1808-16572012000400015
Dufrêne M., Legendre P. 1997. Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecol. Monogr., 67: 345-366. doi:10.1890/0012-9615(1997)067[0345:SAAIST]2.0.CO;2
Dunne J.A., Williams R.J., Martinez N.D. 2002. Food-web structure and network theory: the role of connectance and size. Proc. Natl. Am. Soc., 99: 12917-12922. doi:10.1073/pnas.192407699
Ezcurra C. 1989. Ruellia sanguinea (Acanthaceae) y especies relacionadas en Argentina, Uruguay y sur de Brasil. Darwiniana, 29: 269-287.
Fan Q.H., Flechtmann C.H., Moraes G.J. 2016. Annotated catalogue of Stigmaeidae (Acari: Prostigmata), with a pictorial key to genera. Zootaxa, 4176: 1-199. doi:10.11646/zootaxa.4176.1.1
Ferla N.J., Moraes G.J.D. 2002. Ácaros predadores (Acari) em plantas nativas e cultivadas do Estado do Rio Grande do Sul, Brasil. Rev. Bras. Zool., 19: 1011-1031. doi:10.1590/S0101-81752002000400006
Ferla N.J., Moraes G.J.D. 2003. Biologia de Agistemus floridanus Gonzalez (Acari, Stigmaeidae). Rev. Bras. Zool., 20: 261-264. doi:10.1590/S0101-81752003000200015
Ferla N.J., Silva G. L., Moraes G.J. 2010. Description of a new species of Arrenoseius Wainstein (Acari: Phytoseiidae) from Brazil and a redescription of a similar species from Argentina. Int. J. Acarol., 36: 15-19. doi:10.1080/01647950903490095
Gerson U., Smiley R.L., Ochoa R. 2003. Mites (Acari) for pest control. Blackwell Science Ltd. pp. 540. doi:10.1002/9780470750995
Johann L., Carvalho G.S., Majolo F., Ferla N.J. 2013. Stigmaeid mites (Acari: Stigmaeidae) from vineyards in the state of Rio Grande do Sul, Brazil. Zootaxa, 3701: 238-256. doi:10.11646/zootaxa.3701.2.6
Johann L., Horn T.B., Carvalho G.S., Ferla N.J. 2014. Diversity of mites (Acari) in vineyard agroecosystems (Vitis vinifera) in two viticultural regions of Rio Grande do Sul state, Brazil. Acarologia, 54: 137-154. doi:10.1051/acarologia/20142122
Landeros J., Cerna E., Badii M.H., Varela S., Flores A.E. 2004. Patrón de distribución espacial y fluctuación poblacional de Eutetranychus banksi (McGregor) (Acari: Tetranychidae) y su depredador Euseius mesembrinus (Dean) (Acari: Phytoseiidae) en una huerta de naranjos. Acta Zool. Mex., 20: 147-155.
Lindquist E.E., Krantz G.W., Walter D.E. 2009. Order Mesostigmata. In Krantz G.W., Walter D.E. (Eds). A Manual of Acarology; Texas Tech University Press. p. 124-232.
Lofego A.C., Rezende J.M., Feres R.J.F. 2017. Mite fauna associated with Cecropia pachystachya Trec.(Urticaceae) the importance of the plant as reservoir and dissemination means for predatory mites. Syst. Appl. Acarol., 22: 1780-1794. doi:10.11158/saa.22.10.16
MAPA, Ministério da Agricultura, Pecuária e Abastecimento. 2018. Produtos registrados. [Internet]. [August 3, 2020]. Available online: http://www.agricultura.gov.br
Matioli A.L., Ueckermann E. A., Oliveira C.A.L. 2002. Some stigmaeid and eupalopsellid mites from citrus orchards in Brazil (Acari: Stigmaeidae and Eupalopsellidae). Int. J. Acarol., 28: 99-120. doi:10.1080/01647950208684287
McMurtry J.A., Moraes G. J., Sourassou N.F. 2013. Revision of the lifestyles of phytoseiid mites (Acari: Phytoseiidae) and implications for biological control strategies. Syst. Appl. Acarol., 18: 297-320. doi:10.11158/saa.18.4.1
Mendonça A.L., Lofego A.C., Pott A., Daud R.D., Demite P.R. 2019. Phytoseiidae (Parasitiformes: Mesostigmata) from the Pantanal, Mato Grosso do Sul State, Brazil. Syst. Appl. Acarol., 24:587-612. doi:10.11158/saa.24.4.6
Mineiro J.L.D.C., Sato M.E., Raga A., Arthur V., Moraes G.J., Sarreta F.D.O., Carrijo A. 2006. Diversidade de ácaros (Arachnida: Acari) em Coffea arabica L. cv. Mundo Novo, nos municípios de Jeriquara e Garça, Estado de São Paulo. Biota Neotrop., 6: 1-15. doi:10.1590/S1676-06032006000200012
MMA, Ministério do Meio Ambiente. O bioma Pampa. [Internet]. [August 3, 2020]. Available online: http://www.mma.gov.br/
Moraes G.J., McMurtry J.A., Denmark H.A., Campos C.B. 2004. A revised catalog of the mite family Phytoseiidae. Zootaxa, 434: 1-494. doi:10.11646/zootaxa.434.1.1
Moraes G.J., Flechtmann C.H.W. 2008. Manual de acarologia: acarologia básica e ácaros de plantas cultivadas no Brasil. Ribeirão Preto: Holos Editora. pp. 288.
Niemi G.J., McDonald M.E. 2004. Application of ecological indicators. Ann. Rev. Ecol. Evol. Sci., 35: 89-111. doi:10.1146/annurev.ecolsys.35.112202.130132
Noronha A., Mineiro J.L., Ferla N.J., Castro T.M., Silva G.L., Lofego A.C., Azevedo A.O. 2020. Mites associated with Byrsonima crassifolia (L.) Kunth (Malpighiaceae) in the state of Pará, Brazil. Entomol. Commun., 2: 1-3. doi:10.37486/2675-1305.ec02026
Nuvoloni F.M., Lofego A.C., Rezende J.M., Feres R.J.F. 2015. Phytoseiidae mites associated with Hevea spp. from the Amazon region: a hidden diversity under the canopy of native trees. Syst. Biodiv., 13: 182-206. doi:10.1080/14772000.2014.985344
Onzo A., Hanna R., Sabelis M.W. 2012. The predatory mite Typhlodromalus aripo prefers green-mite induced plant odours from pubescent cassava varieties. Exp. Appl. Acarol., 58: 359-370. doi:10.1007/s10493-012-9595-0
Patrício P.C., Cervi A.C.O. 2005. O gênero Trichilia P. Browne (Meliaceæ) no estado do Paraná, Brasil. Acta Biológica Paranaense, 34: 27-71. doi:10.5380/abpr.v34i0.953
Peel M.C., Finlayson B.L., McMahon T. A. 2007. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sc., 11: 1633-1644. doi:10.5194/hess-11-1633-2007
Pillar V.D., Müller S.C., Castilhos Z.M.S. Jacque A.V.A. 2009. Campos Sulinos: conservação e uso sustentável da biodiversidade. Brasília: Ministério do Meio Ambiente. pp. 403.
R Core Team. 2019. R: A language and environment for statistical computing [Internet]. Vienna, Austria: The R Foundation for Statistical Computing. [August 3, 2020]. Available online: http://https://www.r-project.org/
Rieff G.G., Natal-da-Luz T., Sousa J.P., Wallau M.O., Hahn L., Sá E.L.S. 2016. Collembolans and mites communities as a tool for assessing soil quality: effect of Eucalyptus plantations on soil mesofauna biodiversity. Curr. Sci., 110: 713-719. doi:10.18520/cs/v110/i4/713-719
Rocha M.S., Silva G.L., Oliveira J.S., Freitas E.M., Ferla N.J. 2015. Phytoseiid mites (Acari: Phytoseiidae) in the northern coastal region of the Rio Grande do Sul State, Brazil. Neotrop. Biodiv., 1: 22-35. doi:10.1080/23766808.2015.1052230
Rodrigues E.N.L., Mendonça M.S., Rosado J.L.O., Loeck A.E. 2010. Soil spiders in differing environments: Eucalyptus plantations and grasslands in the Pampa biome, southern Brazil. Rev. Colomb. Entomol., 36: 277-284.
Rodrigues J.K., Furtado I.P., Lofego A.C. 2020. Mites (Arachnida: Acari) associated with plants of family Myrtaceae from the Caatinga Domain, Brazil. Syst. Appl. Acarol., 25: 942-956. doi:10.11158/saa.25.5.14
Streck E.V., Kämpf N., Dalmolin R.S.D., Klamt E., Nascimento P.C., Giasson E., Pinto L.F.S. 2018. Solos do Rio Grande do Sul. Porto Alegre: Emater/RS-Ascar.
Tixier M.S. 2018. Predatory mites (Acari: Phytoseiidae) in agro-ecosystems and conservation biological control: a review and explorative approach for forecasting plant-predatory mite interactions and mite dispersal. Front. Ecol. Evol., 6: 1-21. doi:10.3389/fevo.2018.00192
Vangansbeke D., Nguyen D.T., Audenaert J., Verhoeven R., Deforce K., Gobin B., Tirry L., De Clercq P. 2014. Diet-dependent cannibalism in the omnivorous phytoseiid mite Amblydromalus limonicus. Biol. Contr., 74: 30-35. doi:10.1016/j.biocontrol.2014.03.015
Walter D.E., Lindquist E.E., Smith I.M., Cook D.R., Krantz G.W. 2009. Order Trombidiformes. In Krantz G.W., Walter D.E. (Eds). A Manual of Acarology; Texas Tech University Press. p. 233-420.Walter D.E., Proctor H.C. 2013. Mites: ecology, evolution & behaviour - life at a microscale. New York: Springer. pp 494. doi:10.1007/978-94-007-7164-2



2020-08-09
Date accepted:
2021-01-05
Date published:
2021-01-14
Edited by:
Tixier, Marie Stéphane

This work is licensed under a Creative Commons Attribution 4.0 International License
2021 Toldi, Maicon; de Freitas, Elisete Maria; da Silva, Vinícius Leão; Caumo, Monique; Ferla, Júlia Jantsch; Orlandi, Carla Roberta; Da-Costa, Tairis; Johann, Liana and Ferla, Noeli Juarez
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)