



Phytoseiid mites of Mayotte Island (Acari: Mesostigmata)
Kreiter, Serge  1
; Abo-Shnaf, Reham I.A.
2
and Payet, Rose-My
3
1
; Abo-Shnaf, Reham I.A.
2
and Payet, Rose-My
3
1✉ Montpellier SupAgro, UMR CBGP INRAE/ IRD/ CIRAD/ SupAgro, Université de Montpellier, 755 Avenue du Campus Agropolis (Baillarguet), CS 30016, 34988 Montferrier-sur-Lez cedex, France.
2Plant Protection Research Institute (PPRI), Agricultural Research Centre (ARC), 7 Nadi El-Seid Street, Dokii, 12611 Giza, Egypt.
3CIRAD, Université de Montpellier, Unité Hortsys, Station de Bassin-Plat, 97410 Saint-Pierre, La Réunion, France.
2020 - Volume: 60 Issue: 3 pages: 622-642
https://doi.org/10.24349/acarologia/20204391Original research
Keywords
Abstract
Introduction
Mites of the family Phytoseiidae (Acari: Mesostigmata) are well-known for their predatory behaviour on phytophagous mites and small insects on cultivated plants and wild vegetation. Some of them are used to control pest organisms especially in protected crops and to some extent in open fields all around the world (McMurtry and Croft 1997; McMurtry et al. 2013). This family is widespread around the world, presents on all continents (except Antarctica). It presently consists of more than 2,500 valid species belonging to 94 genera and three subfamilies (Demite et al. 2020). Biodiversity surveys in poorly investigated areas is still an urgent needed and might result in the discovery of additional species potentially useful for biological control as well as having more information on the biodiversity of these areas (Kreiter et al. 2018a, b, c, 2020a, b, c). The more interesting area are probably those with a high level of biodiversity. Most of the Indian Ocean constitutes one of the world biodiversity hotspots, a concept defined by Myers (1988) in order to identify the most immediately important areas for biodiversity conservation. The common characteristics of these hotspots is that they hold high endemism levels and have lost at least 70% of their original natural vegetation (Myers et al. 2000). Knowledge of the phytoseiid diversity in these high interest areas in the context of global climate changes may contribute to identify potential biological control agent (BCA) and future establishment of conservation programs. Located in the Indian Ocean at 300 km from the northern coast of Madagascar, Mayotte Island (Maore in Shikomori language) is one of the four main islands constituting Comoros Archipelago, with Anjouan (Ndzuwani or Nzwani in Shikomori, and Johanna or Anjouane in Arabic), Mohéli (Mwali in Shikomori) and Grande Comore (Ngazidja in Shikomori). Only one phytoseiid species had been reported from this island long time ago, namely Phytoseius mayottae Schicha (Schicha 1984). The objective of this paper is to report the phytoseiid species found in a new survey conducted in November 2018 in Mayotte Island.
Material and methods
The survey took place in Mayotte in the second half of November 2018. Plant inhabiting mites were collected from cultivated and uncultivated plants in few locations in the centre of the island. Mites were directly collected on leaves with a fine brush with or without a pocket lens or a stereo-microscope when available (large leaf and herbaceous plants) or by beating the plants (mainly shrubs and trees with very small or spiny leaves) and collecting the mites in a black plastic rectangular saucer 45 x 30 cm (Ref. STR 45, BHR, 71370 Saint-Germain-du-Plain, France). The mites were then transferred with a fine brush into small plastic vials containing 1.5 ml of 70% ethanol. All mites were mounted on slides using Hoyer's medium and they were identified using a phase and interferential contrast microscope (DMLB, Leica Microsystèmes SAS, Nanterre, France). Morphological characters of specimens were measured using a graded eyepiece (Leica, see above). Chant and McMurtry's (1994, 2007) concepts of the taxonomy of the family Phytoseiidae for identification and the world catalogue database of Demite et al. (2014, 2020) for distribution and information on descriptions and re-descriptions were used. The setal nomenclature system adopted was that of Lindquist & Evans (1965) and Lindquist (1994) as adapted by Rowell et al. (1978), and Chant & Yoshida-Shaul (1992) for the dorsum and by Chant & Yoshida-Shaul (1991) for the venter. The notation for solenostomes and poroids is based on Athias-Henriot (1975). Numbers of teeth on the fixed and movable cheliceral digits do not include the respective apical teeth on apical hook. Setae not referred to in the results section should be considered as absent. All measurements are given in micrometres (µm) and presented with the mean in bold followed by the range in parenthesis.
Specimens of each species are deposited in the mite collections of Montpellier SupAgro conserved in UMR CBGP INRAE/IRD/CIRAD/SupAgro/University of Montpellier. Specimens collected in fields in Mayotte within these surveys were all identified. Only few single males or immatures collected during this study are not taken into account. The following abbreviations are used in this paper for morphological characters: dsl = dorsal shield length just above j1 to just below J5; dsw = dorsal shield width at the level of s4; Z4 ser., Z5 ser. = Z4, Z5 serrated (if Z4 and Z5 without ser. = not serrated); gensl = genital shield length; gensw post. cor. = genital shield width posteriorly; lisl = Largest inguinal sigilla (= ''metapodal plate'') length; lisw = Largest inguinal sigilla (= ''metapodal plate'') width; sisl = smallest inguinal sigilla (= ''metapodal plate'') length; sisw = smallest inguinal sigilla (= ''metapodal plate'') width; vsl = ventrianal shield length; gv3 – gv3 = distance between solenostomes gv3 on the ventrianal shield; vsw ZV2 & vsw anus = ventrianal shield width at ZV2 level and at paranal setae level; scl: calyx length; scw = calyx widest width; Fdl = fixed digit length; Mdl = movable digit length; No teeth Fd = number of teeth on the fixed digit; No teeth Md = number of teeth on the movable digit; Shaft = length of the shaft of spermatodactyl; toe = length of the toe; BCA = Biological control agent; aasl = altitude above sea level. The following abbreviations are used in this paper for institutions: CBGP = Centre de Biologie pour la Gestion des Populations; CIRAD = Centre International de Recherche Agronomique pour le Développement; INRAE = Institut National de Recherche pour l'Agriculture, l'Alimentation et l'Environnement; IRD = Institut de Recherche pour le Développement; MSA = Montpellier SupAgro, France; UMR = Unité Mixte de Recherche; UR = Unité de Recherche.
Results and discussion
A total of 18 species are found, sixteen presented thereafter.
Subfamily Amblyseiinae Muma
Amblyseiinae Muma 1961: 273.
Tribe Neoseiulini Chant & McMurtry
Neoseiulini Chant & McMurtry 2003a: 6.
Genus Neoseiulus Hughes
Neoseiulus Hughes 1948: 141.
Neoseiulus barkeri Hughes
Neoseiulus barkeri Hughes 1948: 141; Ryu et al. 2001: 8; Chant & McMurtry 2003a: 35, 2007: 25; Moraes et al. 1986:70, 2004: 104.
Typhlodromus (Neoseiulus) barkeri, Nesbitt 1951: 35.
Typhlodromus (Typhlodromus) barkeri, Chant 1959: 63.
Amblyseius barkeri, Athias-Henriot 1961: 440; Moraes et al. 1989: 95.
Typhlodromus (Amblyseius) barkeri, Hughes 1961: 222.
Typhlodromus barkeri, Hirschmann 1962: 5.
Amblyseius (Amblyseius) barkeri, van der Merwe 1968: 112.
Amblyseius mckenziei Schuster & Pritchard 1963: 268 (synonymy according to Ragusa & Athias-Henriot 1983).
Amblyseius usitatus van der Merwe 1965: 71 (synonymy according to Ueckermann & Loots 1988).
Amblyseius oahuensis Prasad 1968: 1518 (synonymy according to Ragusa & Athias-Henriot 1983).
Amblyseius picketti Specht 1968: 681 (synonymy according to Ragusa & Athias-Henriot 1983).
Amblyseius mycophilus Karg, 1970: 290 (synonymy according to Ragusa & Athias-Henriot 1983).
Amblyseius masiaka Blommers & Chazeau 1974: 308 (synonymy according to Ueckermann & Loots 1988).
This species belongs to the barkeri species group of the genus Neoseiulus and to the barkeri species subgroup (Chant and McMurtry 2003a). Neoseiulus barkeri has a worldwide distribution (Moraes et al. 2004; Demite et al. 2020). Various studies have shown its ability to control Frankliniella occidentalis Pergande (Rodriguez-Reina et al. 1992), Thrips tabaci (Lindeman) (Broodsgaard and Stengaard Hansen 1992) and Tetranychus urticae Koch on cucumbers (Fan and Petitt 1994b). Fan and Petitt (1994a) showed that augmentative releases of N. barkeri provided also control of the broad mite, Polyphagotarsonemus latus (Banks), on peppers. Neoseiulus barkeri constitutes a potential BCA for several crops, especially in vegetables greenhouses. This species has been mentioned by Quilici et al. (2000) and Kreiter et al. (2020c) in La Réunion Island and more recently from Rodrigues by Kreiter and Abo-Shnaf (2020a).
World distribution: it has a worldwide distribution in all continents, in more than 50 countries (Demite et al. 2020).
Specimens examined: a single ♀ specimen collected during this study. L'Abattoir, City Center (15 m aasl, 12°47'18'' S, 45°16'21'' E), 1 ♀ on Hibiscus rosa-sinensis L. (Malvaceae) with eriophyid mite galls, 14/XI/2018.
Remarks: measurements of characters of female from Mayotte are only slightly different from female specimens from other countries, especially La Réunion Island. Comparisons with N. barkeri measurements of female and male specimens of various origins in Beaulieu and Beard (2018) shows shorter dimensions of all characters of Mayotte specimens. These authors already mentioned the shorter dimensions of dorsal setae of African female and male specimens (lower part of observed ranges) compared to their measurements of type and additional materials (Beaulieu and Beard 2018).
Neoseiulus teke (Pritchard and Baker)
Amblyseius (Amblyseius) teke Pritchard & Baker 1962: 239.
Amblyseius teke, Meyer & Rodrigues 1966: 30; Moraes et al. 1989a: 83, 1989b: 97.
Neoseiulus teke, Moraes et al. 1986: 98, 2004: 147; Chant & McMurtry 2003a: 37, 2007: 31.
Amblyseius (Amblyseius) bibens Blommers 1973: 111 (synonymy according to Ueckermann & Loots 1988).
Neoseiulus teke belongs to the barkeri species group and the womersleyi species subgroup (Chant and McMurtry 2003a). This species is found in sub-Saharan Africa often associated with Mononychellus tanajoa (Bondar), the cassava green mite (CGM). It has been studied for its potential use as a BCA against the CGM. Nwilene and Nachman (1996) studied its reproduction characteristics on M. tanajoa. It was more efficient than Iphiseius degenerans (Berlese), but seems not efficient enough in field conditions (Nwilene and Nachman 1996). Quilici et al. (2000) collected this species in La Réunion Island and it was reported recently by Kreiter et al. (2020c).
World distribution: Burundi, DR Congo, Ghana, Kenya, Malawi, Mozambique, La Réunion Island, Rwanda, Sierra Leone, South Africa, Tanzania, Zimbabwe.
Specimens examined: 4 specimens in total, 3 ♀♀ + 1 ♂. L'Abattoir, City Center (15 m aasl, 12°47'18'' S, 45°16'21'' E), 1 ♀ + 1 ♂ on Hibiscus rosa-sinensis L. (Malvaceae) with eriophyid mite galls and 2 ♀♀ on Carica papaya L. (Caricaceae), 27/XI/2018.
Remarks: measurements of morphological characters of N. teke female and male specimens from Mayotte Island (Tables 1 and 2) are very close to those specimens from neighbouring countries, especially from specimens of La Réunion Island (Kreiter et al. 2020c) and various countries in Africa, except for the holotype (Zannou et al. 2006) and specimens from South Africa which are larger (van der Merwe 1965).



Tribe Kampimodromini Kolodochka
Kampimodromini Kolodochka 1998: 59.
Subtribe Paraphytoseiina Chant & McMurtry
Paraphytoseiina Chant & McMurtry 2003b: 211.
Genus Paraphytoseius Swirskii & Shechter
Paraphytoseius Swirski & Shechter 1961: 113.
Paraphytoseius horrifer (Pritchard & Baker)
Amblyseius (Ptenoseius) horrifer Pritchard & Baker 1962: 295.
Amblyseius horrifer, Meyer & Rodrigues 1966: 30.
Amblyseius (Paraphytoseius) horrifer, van der Merwe 1968: 169.
Proprioseius (Paraphytoseius) horrifer, Karg 1983: 302.
Paraphytoseius horrifer, Moraes et al. 1986: 105, 2004: 152; Beard 2001: 84; Chant & McMurtry 2003a: 37, 2007: 53.
In all of our specimens of Paraphytoseius, setae S5 are absent. So according to Chant and McMurtry (2003b), they belong to the orientalis species group. According also to these previous authors, and Moraes et al. (2007), we consider that P. horrifer and P. orientalis (Narayanan, Kaur & Ghai) are different valid species. Our specimens with longer setae s4, Z4, Z5, and with no distinctly short, thick, spatulate macroseta on genu I belongs to the former species. This species is widely distributed in Sub-Saharan Africa and Madagascar. The biology of P. horrifer remains totally unknown. It was recently recorded for the first time from several countries: Mauritius (Kreiter and Abo-Shnaf 2020b), La Réunion (Kreiter et al. 2020c), Rodrigues (Kreiter and Abo-Shnaf 2020a) and Vietnam (Kreiter et al. 2020b).
World distribution: Benin, DR Congo, Ghana, India, Kenya, La Réunion Island, Madagascar Island, Malawi, Mozambique, Senegal, South Africa, Uganda.
Specimens examined: 11 specimens in total, 7 ♀♀ + 4 ♂. Coconi, Maison de L'Office National des Forêts (156 m aasl, 12°50'1'' S, 45°8'5'' E), 1 ♀ + 1 ♂ on Cananga odorata L. (Annonaceae) and 4 ♀♀ + 1 ♂ on an unknown host plant, 24/XI/2018; Miréréni (356 m aasl, 12°47'45'' S, 45°9'28'' E), 1 ♀ + 2 ♂♂ on Rubus alceifolius Poiret (Rosaceae); Coconi, Lycée Agricole (189 m aasl, 12°50'7'' S, 45°8'11'' E), 1 ♀ on R. alceifolius, 26/XI/2018.
Remarks: morphological and morphometric characters and all measurements fit well with those provided by Kreiter et al. (2020b, c). This species described from Africa (Pritchard and Baker 1962) was first mentioned in the Indian Ocean from La Réunion Island (Kreiter et al. 2020c), but seems to be present in several other investigated islands (Kreiter and Abo-Shnaf 2020a, b).
Paraphytoseius orientalis (Narayanan, Kaur & Ghai)
Typhlodromus (Amblyseius) orientalis Narayanan, Kaur & Ghai 1960: 394.
Paraphytoseius orientalis, Moraes et al. 1986: 105, 2004: 162; Chant & McMurtry 2003b: 220, 2007: 53.
Amblyseius ipomeai El-Banhawy 1984: 126 (synonymy according to Chant & McMurtry 2003b).
Paraphytoseius multidentatus Swirski & Shechter 1961: 114 (synonymy according to Matthysse & Denmark 1981 in Denmark et al. 1999).
Paraphytoseius narayanani Ehara 1967: 67 (synonymy according to Chant & McMurtry 2003b).
This species belongs to the orientalis species group (Chant and McMurtry 2003b), but according to these authors and Moraes et al. (2007), specimens with shorter setae s4, Z4 and Z5, and having a distinctly short, thick, spatulate macroseta on genu I belong to the species P. orientalis. This species is widely distributed in tropical and subtropical areas in South America, Africa and Asia. It belongs to a genus included in the large polyphagous generalist group named type III phytoseiid mites (McMurtry and Croft 1997; McMurtry et al. 2013). Navasero and Navasero (2016) studied the life history of P. orientalis on the broad mite (P. latus) as prey and reported high predation rates when eggs of P. latus were offered as food, suggesting good potential for the control of this pest. This species was also collected in Mauritius (Kreiter et al. 2018a; Ferragut and Baumann 2019; Kreiter and Abo-Shnaf 2020b), La Réunion (Kreiter et al. 2020c) and Vietnam (Kreiter et al. 2020b).
World distribution: Argentina, Brazil, Burundi, India, Japan, Kenya, La Réunion Island, Madagascar Island, Martinique Island, Mauritius Island, Mozambique, Rwanda.
Specimens examined: 3 ♀♀ specimens in total. Coconi, Lycée Agricole (189 m aasl, 12°50'7'' S, 45°8'11'' E), 3 ♀♀ on Capsicum annuum L. (Solanaceae), 26/XI/2018.
Remarks: this species was reported by Kreiter et al. (2018a, 2020b, c) and by Ferragut and Baumann (2019) from various places. Morphological and morphometric characters and all measurements fit well with those given by Ferragut and Baumann (2019) and Kreiter et al. (2020b, c). It is described from Asia (Narayanan et al. 1960) and present in Vietnam (Kreiter et al. 2020b). Paraphytoseius orientalis seems to be more common than P. horrifer in Mauritius Island (Kreiter and Abo-Shnaf 2020b), whereas the latter is more abundant in Mayotte Island.
Tribe Typhlodromipsini Chant & McMurtry
Typhlodromipsini Chant & McMurtry 2005c: 318.
Genus Typhlodromips De Leon
Typhlodromips 2007: 55.
Typhlodromips shi (Pritchard & Baker)
Amblyseius (Amblyseius) shi Pritchard & Baker 1962: 252.
Typhlodromips shi, Moraes et al. 1986: 147, 2004: 224; Chant & McMurtry 2005c: 327, 2007: 63.
Typhlodromips ivoloinae (Blommers 1974): 146 (synonymy according to Ueckermann & Loots 1988).
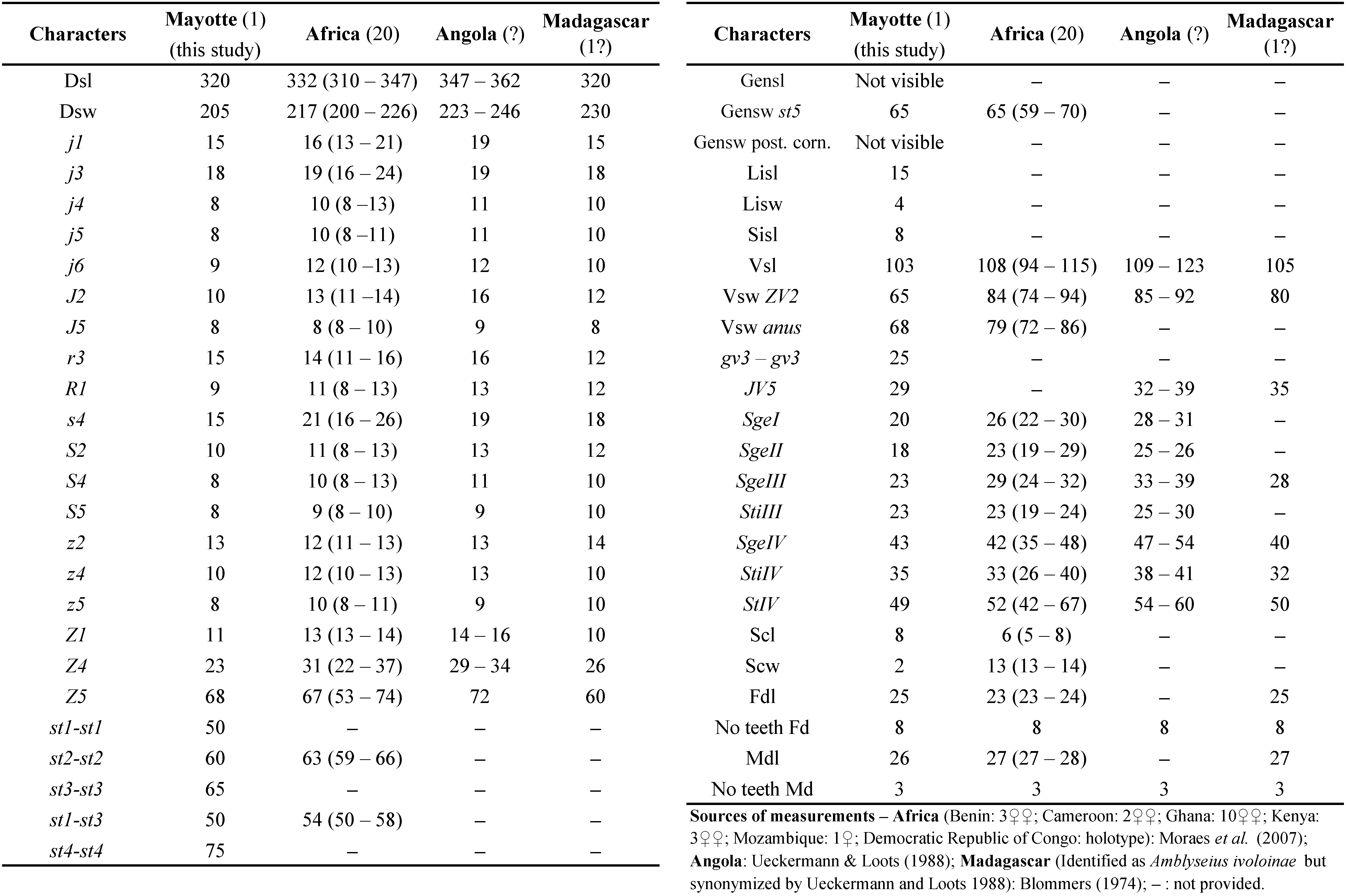
This species belongs to the culmulus species group of the genus Typhlodromips with nine other species. It was described under the name ivoloinae by Blommers (1974) from Madagascar on Citrus lemon (L.) Burman (Rutaceae). Mayotte Island is approximately 340 km away from Madagascar coasts and it is not surprising to find this species on a close island. The species was not reported from Mascareignes Archipelago. Its biology is totally unknown.
World distribution: Angola, Benin, Cameroon, DR Congo, Ghana, Kenya, Madagascar, Malawi, Mozambique, Nigeria, Sierra Leone.
Specimens examined: a single ♀ collected during this study. Coconi, Lycée Agricole (189 m aasl, 12°50'7'' S, 45°8'11'' E), 1 ♀ on Carica papaya L. (Caricaceae), 26/XI/2018.
Remarks: morphological and morphometric characters and all measurements fit well with those provided in the literature as indicated in (Table 3). This species recorded in several countries of Africa and is also present in Madagascar. Mayotte is consequently the second island of Indian Ocean known for hosting this species.
Tribe Amblyseiini Muma
Amblyseiinae Muma 1961: 273.
Amblyseiini Muma, Wainstein 1962: 26.
Subtribe Amblyseiina Muma
Amblyseiina Muma, Chant & McMurtry 2004: 179.
Genus Amblyseius Berlese
Amblyseius Berlese 1914: 143.
Amblyseius largoensis (Muma)
Amblyseiopsis largoensis Muma 1955: 266.
Typhlodromus (Amblyseius) largoensis, Chant 1959: 96.
Amblyseius (Amblyseialus) largoensis, Muma 1961: 287.
Typhlodromus largoensis, Hirschmann 1962: 2.
Amblyseius (Amblyseius) largoensis, Ehara 1966: 22.
Amblyseius largoensis, Swirski & Golan 1967: 225; Moraes et al. 1986: 17, 2004: 33; Chant & McMurtry 2004: 208, 2007: 78.
Amblyseius magnolia Muma 1961: 289 (synonymy according to Denmark & Evans 2011).
Amblyseius sakalava Blommers 1976: 96 (synonymy according to Ueckermann & Loots 1988).
Amblyseius amtalaensis Gupta 1977: 53 (synonymy according to Gupta 1986).
Amblyseius largoensis belongs to the largoensis species group and the largoensis species subgroup. It is widespread in all tropical and subtropical regions of the world and was the most abundant species collected by Moraes et al. (2000) in French Caribbean Islands and as a potential BCA of Raoiella indica Hirst in La Réunion Island (Moraes et al. 2012). Using morphometric analyses of 36 characters, molecular analyses and crossing tests, Navia et al. (2014) studied specimens collected in Brazil, La Réunion Island and Trinidad and Tobago to determine whether A. largoensis populations from different geographic origins belong to the same taxonomic entity. Though differences in the lengths of some setae were observed, molecular analyses and crossing experiments indicated that populations from Indian Ocean and America were conspecific. This species was previously recorded from Mauritius Island by Ferragut and Baumann (2019) and Kreiter and Abo-Shnaf (2020b), and Rodrigues Island by Kreiter and Abo-Shnaf (2020a).
World distribution: this species is widely distributed in the tropical and subtropical regions of Africa, America, Asia and the Pacific Islands.
Specimens examined: 17 specimens in total, 8 ♀♀, 4 ♂♂ and 5 im. L'Abattoir, Dziani lake (23 m aasl, 12°46'14'' S, 45°17'18'' E), 1 ♂ and 2 im. on Phoenix dactyliphera L. (Arecaceae) and 1 ♂ on Artocarpus altilis (Parkinson) Fosberg (Moaceae) 27/XI/2018; L'Abattoir, City Center (15 m aasl, 12°47'18'' S, 45°16'21'' E), 2 ♀♀ and 1 im. on Mangifera indica L. (Anacardiaceae), 3 ♀♀ and 2 ♂♂ on Hibiscus rosa-sinensis (Malvaceae) with eriophid mite galls, and 3 ♀♀ and 2 im. on Cordia sebestena (Boraginaceae), 27/XI/2018.
Remarks: morphological and morphometric characters and all measurements of Mayotte Island specimens fit well with those given in Zannou et al. (2007) for specimens from Africa, Navia et al. (2014) for specimens from Brazil, La Réunion and Trinidad & Tobago and Ferragut and Baumann (2019) for specimens from Mauritius. This is the third more common species of our samplings.
Amblyseius parasundi Blommers
Amblyseius (Proprioseiopsis) parasundi Blommers 1974: 144.
Amblyseius parasundi, Moraes et al. 1986: 27, 2004: 46.
Amblyseius (Amblyseius) parasundi, Denmark & Muma 1989: 19.
This species have no setae Z1 and consequently belongs to the sundi species group and having the spermatheca elongate, tub-like, belongs to the sundi species subgroup. Despite its high population on fruit trees in Madagascar and preying on tetranychid mites (Blommers and Gutierrez 1975), its biology is totally unknown.
World distribution: Madagascar Island.
Specimens examined: 29 specimens in total, 24 ♀♀, 4 ♂♂ and 1 im. Coconi, Maison de L'Office National des Forêts (156 m aasl, 12°50'1'' S, 45°8'5'' E), 1 ♀ on Persea americana Miller (Lauraceae) and 2 ♀♀ + 1 ♂ on Terminalia catappa L. (Combretaceae), 24/XI/2018; Miréréni (356 m aasl, 12°47'45'' S, 45°9'28'' E), 1 ♀ on Calophyllum inophyllum Poiret (Calophyllaceae) and 1 ♀ on Pteridium aqualinum (L.) Kuhn (Dennstaedtiaceae), 24/XI/2018; Combani, gîte du Mont Combani (437 m aasl, 12°48'23'' S, 45°9'17'' E), 2 ♀♀ on Artocarpus heterophyllus Lamarck (Moraceae), 1 ♀ on Malvaviscus arboreus Cavanilles (Malvaceae), 1 ♀ and 1 ♂ on Cananga odorata (Lamarck) Hook & Thomson (Annonaceae), 3 ♀♀ on Psidium guajava L. (Myrtaceae), 1 ♂ on Cocos nucifera L. (Arecaceae), 2 ♀♀ on Artocarpus altilis (Parkinson) Fosberg (Moraceae), 1 ♀ on P. americana and 2 ♀♀ on Psidium cattleianum Afzelius ex. Sabine (Myrtaceae), 25/XI/2018; Coconi, Lycée Agricole (189 m aasl, 12°50'7'' S, 45°8'11'' E), 1 ♀ on A. altilis, 1 ♀ on Mangifera indica L. (Anacardiaceae), 1 ♀ on Syngonium podophyllum Schott (Araceae) and 1 ♀ on Piper nigrum L. (Piperaceae), 26/XI/2018; Combani, grower farm (104 m aasl, 12°47'14'' S, 45°7'57'' E), 2 ♀♀ on C. odorata and 1 ♀ and 1 im. on Theobroma cacao L. (Malvaceae), 26/XI/2018; L'Abattoir, Dziani lake (23 m aasl, 12°46'14'' S, 45°17'18'' E), 1 ♂ on A. altilis, 27/XI/2018.
Remarks: morphological and morphometric characters and all measurements (Table 4) fit quite well with few measurements from the other studies (Table 4) except some setae which are 10–20% longer (Z5, JV5, SgeIV, StiIV and StIV) or 10–40% shorter (j3, r3) in Mayotte specimens. This is the second more common species of our samplings. Amblyseius sundi is reported by Blommers (1974) as being a thelytokous species in mass-rearing and field collected specimens and similar information is also mentioned by Denmark and Muma (1989). Four males were however collected during our study.

Amblyseius tamatavensis Blommers
Amblyseius tamatavensis Blommers 1974: 144; Moraes et al. 1986: 31, 2004: 52; Denmark & Muma 1989: 13; Chant & McMurtry 2004: 203, 2007: 81; Ehara & Amano 2004: 17.
Amblyseius maai Tseng 1976: 123 (synonymy according to Denmark & Muma 1989).
Amblyseius aegyptiacus Denmark & Matthysse in Matthysse & Denmark 1981: 343 (synonymy according to Denmark & Muma 1989).
Amblyseius (Amblyseius) tamatavensis, Ehara 2002: 33; Ehara & Amano 2002: 322.
Amblyseius tamatavensis belongs to the obtusus species group and the aerialis species subgroup which contains 46 species (Chant and McMurtry 2004). It seems to fit the functional type III-b (generalist predators living on glabrous leaves) group defined by McMurtry et al. (2013). Cavalcante et al. (2017) reported this species as a promising natural enemy of Bemisia tabaci (Gennadius). Experimental releases of this predator on caged plants in a screenhouse caused the reduction of the density of B. tabaci on pepper plants by up to 60–80 % (Massaro and Moraes 2019). It can be easily produced in large numbers (Massaro et al. 2018) when fed with astigmatine mites, which could allow the mass production for augmentative biological control. This species is reported in tropical areas from over 20 countries around the world (Africa, Asia, America and Oceania). It was recorded from La Réunion Island (Quilici et al. 2000; Kreiter et al. 2020c), from Rodrigues Island (Kreiter and Abo-Shnaf 2020a) and from Mauritius Island (Ferragut and Baumann 2019; Kreiter and Abo-Shnaf 2020b).
World distribution: this species was described from Madagascar, but is actually widely distributed in the tropical and subtropical regions of Africa, America, Asia and the Pacific Islands.
Specimens examined: 4 specimens in total, 3 ♀♀ and 1 ♂. Combani, grower farm (104 m aasl, 12°47'14'' S, 45°7'57'' E), 3 ♀♀ on Capsicum annuum L. (Solanaceae), 26/XI/2018 and 1 ♂ on Citrus sinensis (L.) Osbeck (Rutaceae), 26/XI/2018.
Remarks: this species was described from Madagascar (Blommers 1974), then was mentioned in the Indian Ocean from La Réunion Island (Quilici et al. 2000). Morphological and morphometric characters and all measurements of our specimens fit well with those provided in Blommers (1974) for specimens from Madagascar, Ferragut and Baumann (2019) for specimens from Mauritius Island and Kreiter et al. (2020c) for specimens from La Réunion Island.
Subtribe Proprioseiopsina Chant & McMurtry
Proprioseiopsina Chant & McMurtry, 2004: 219.
Genus Proprioseiopsis Muma
Proprioseiopsis Muma 1961: 277.
Proprioseiopsis ovatus (Garman)
Amblyseiopsis ovatus Garman 1958: 78.
Typhlodromus (Amblyseius) ovatus, Chant 1959: 90.
Amblyseiulus ovatus, Muma 1961: 278.
Typhlodromus ovatus, Hirschmann 1962: 2.
Proprioseiopsis ovatus, Moraes et al. 1986: 121, 2004: 184; Chant & McMurtry 2005a: 15, 2007: 89.
Proprioseiopsis (Proprioseiopsis) ovatus, Karg 1989: 208.
Proprioseiopsis cannaensis (Muma 1962: 4) (synonymy according to Denmark & Evans 2011).
Proprioseiopsis peltatus (van der Merwe: 1968: 119) (synonymy according to Tseng 1983).
Proprioseiopsis hudsonianus (Chant & Hansell 1971: 723) (synonymy according to Denmark & Evans 2011).
Proprioseiopsis parapeltatus (Wu & Chou, 1981: 274) (synonymy according to Tseng 1983).
Proprioseiopsis antonelli Congdon 2002: 15 (synonymy according to Denmark & Evans 2011).
Proprioseiopsis ovatus belongs to the belizensis species group as genu I have no macrosetae. As the spermatheca of that species is saccular, it belongs to the belizensis species subgroup (Chant and McMurtry 2005a). This species is known from Guadeloupe, Marie-Galante and Martinique (Kreiter and Moraes 1997; Moraes et al. 2000; Mailloux et al. 2010; Kreiter et al. 2018c). It was found in very large number only during a previous study on companion plant in Guadeloupe (Mailloux et al. 2010) and in a recent study in La Réunion (Le Bellec, unpub. data). In other habitats, it seems to be rare. Similar to P. mexicanus (Garman), P. ovatus seems to be abundant on weeds in the lower vegetation. Denmark and Evans (2011) indicated that this species is associated with Oligonychus pratensis (Banks) and Brevipalpus spp. It was also found in association with Tetranychus evansi Baker and Pritchard (Furtado et al. 2014), but mentioned as poor predator of that species. Despite this information, the biology of P. ovatus remains unknown.
World distribution: Argentina, Brazil, Colombia, Costa Rica, Cuba, Ecuador, Egypt, Ghana, Hawaii, Honduras, Japan, Malaysia, Martinique Island, Mozambique, Peru, Phiippines, Puerto Rico, La Réunion Island, Saudi Arabia, Sierra Leone, South Africa, Spain, Sri Lanka, Taiwan, Thailand, Turkey, USA, Venezuela.
Specimens examined: a single ♀ collected during this study. Combani, gîte du Mont Combani (437 m aasl, 12°48'23'' S, 45°9'17'' E), 1 ♀ on Hydrangea aspera Buch.-Ham. ex D. Don (Hydrangeaceae), 25/XI/2018.
Remarks: morphological and morphometric characters and all measurements of our single specimen in perfect shape fit well measurements of Kreiter et al. (2020c) and other measurements of the literature mentioned by these authors for specimens from La Réunion and other parts of the world.
Tribe Euseiini Chant & McMurtry
Euseiini Chant & McMurtry 2005b: 191.
Subtribe Euseiina Chant & McMurtry
Euseiina Chant & McMurtry 2005b: 209.
Genus Moraeseius Chant & McMurtry
Moraeseius Chant & McMurtry 2005b: 209.
Moraeseius papayana (van der Merwe)
Amblyseius (Amblyseius) papayana van der Merwe 1965: 57.
Amblyseius (Proprioseiospsis) papayana, van der Merwe 1968: 161.
Euseius papayana, Moraes et al. 1986: 46, 2001: 46, 2004: 78.
Moraeseius papayana, Chant & McMurtry 2005b: 216, 2007: 123.
Moraeseius papayana was first placed in the genus Amblyseius, then assigned to the genus Euseius before being erected as a new genus, Moraeseius by Chant and McMurtry (2005b). It was described from Nelspruit, Transvaal, South Africa on Carica papaya L. This is the first record of this species outside the African continent. The biology of this species is totally unknown.
World distribution: Kenya, Mozambique, South Africa.
Specimens examined: 2 ♀♀ specimens in total. Combani, gîte du Mont Combani (437 m aasl, 12°48'23'' S, 45°9'17'' E), 2 ♀♀ on Carica papaya L. (Caricaceae), 25/XI/2018.
Remarks: morphological and morphometric characters and all measurements of our specimens (Table 5) fit well with those of van der Merwe (1965) in the original description for specimens from South Africa and of Moraes et al. (2001) for specimens from Kenya.

Genus Euseius Wainstein
Amblyseius (Amblyseius) section Euseius Wainstein 1962: 15.
Euseius De Leon 1966: 86.
Euseius ovaloides (Blommers)
Amblyseius (Amblyseius) ovaloides Blommers 1974: 147.
Euseius ovaloides, Moraes et al. 1986: 51, 2004: 78; Chant & McMurtry 2005a: 215, 2007: 121.
Euseius ovaloides was described by Blommers (1974) from specimens collected on Citrus hystrix de Candolle (Rutaceae) and Persea americana Miller (Lauraceae) in Madagascar. Like all Euseius species, this species belongs to the type IV (polliniphagous generalist predators) of McMurtry and Croft (1997) and McMurtry et al. (2013). The species had been occasionally recorded from Madagascar (Blommers 1974), Papua-New Guinea (Schicha and Gutierrez 1985), Seychelles (Schicha 1987), La Réunion Island, (Quilici et al. 1997, 2000, Kreiter et al. 2020c), Guadeloupe, Martinique and Marie-Galante (Moraes et al. 2000; Kreiter et al. 2006) on various plants, though its biology remains unknown. It is suspected to be a poor predator of tetranychid mites (Gutierrez and Etienne 1986), but can be considered as a potential predator of thrips and whiteflies. This is one of the most common species on La Réunion Island (Kreiter et al. 2020c).
World distribution: Guadeloupe, Madagascar Island, Marie-Galante, Martinique, Papua New Guinea, La Réunion Island, Seychelles Archipelago.
Specimens examined: 4 ♀♀ specimens in total. L'Abattoir, City Center (15 m aasl, 12°47'18'' S, 45°16'21'' E), 4 ♀♀ Carica papaya L. (Caricaceae), 27/XI/2018.
Remarks: this species was recently reported from Vietnam (Kreiter et al. 2020b). Morphological and morphometric characters and all measurements of our specimens fit well with measurements in Kreiter et al. (2020b). This species was the second more collected species in our study concerning Mauritius (Kreiter & abo-Shnaf 2020b) Phytoseiidae after A. herbicolus and it was also very common in La Réunion (Kreiter et al. 2020c), but less common in Mayotte Island.
Subfamily Phytoseiinae Berlese
Phytoseiini Berlese 1913: 3.
Phytoseiinae Vitzthum 1941: 767.
Genus Phytoseius Ribaga
Phytoseius Ribaga 1904: 177.
Phytoseius haroldi Ueckermann & Kreiter
Phytoseius haroldi Ueckermann & Kreiter in Kreiter et al. 2002: 339; Chant & McMurtry 2007: 129.
This species belongs to the horridus species group as setae J2 and R1 are absent (Chant and McMurtry 1994). This species was described by Ueckermann and Kreiter in Kreiter et al. (2002) from La Réunion Island. It was abundant in low vegetation in a study of companion plants in citrus orchard in La Réunion Island (Kreiter et al. 2020c). It seems that this species prefers low plants, but this observation has to be confirmed in further studies. The biology of this species remains also totally unknown. This species was recently reported in Mauritius (Ferragut and Baumann 2019; Kreiter and Abo-Shnaf 2020b) and in Rodrigues Island (Kreiter and Abo-Shnaf 2020a).
World distribution: La Réunion Island, Mauritius Island, Rodrigues Island.
Specimens examined: a single ♀ collected during this study. L'Abattoir, Dziani lake (23 m aasl, 12°46'14'' S, 45°17'18'' E), 1 ♀ on Anacardium occidentale L. (Anacardiaceae), 27/XI/2018
Remarks: morphological and morphometric characters and all measurements of our specimens fit well with those of the original description in Kreiter et al. (2002) concerning specimens from La Réunion Island, Ferragut and Baumann (2019) for specimens from Mauritius, Kreiter et al. (2020c) for additional specimens from La Réunion Island and Kreiter and Abo-Shnaf (2020a) for specimens from Rodrigues Island.
Subfamily Typhlodrominae Wainstein
Typhlodromini Wainstein 1962: 26.
Typhlodrominae Chant & McMurtry 1994: 235.
Tribe Chanteiini Chant & McMurtry
Chanteiini Chant & McMurtry 1994: 237, 2007: 132.
Genus Chanteius Wainstein
Chanteius Wainstein 1962: 19.
Chanteius contiguus (Chant)
Typhlodromus (Typhlodromus) contiguus Chant, 1959: 29.
Typhlodromus (Diadromus) contiguus, Athias-Henriot 1960: 62.
Typhloseiopsis contiguus, Muma 1961: 294.
Chanteius (Chanteius) contiguus, Wainstein 1962: 9.
Typhlodromus contiguus, Hirshmann 1962: 2.
Typhlodromus (Typhloseiopsis) contiguus, Pritchard & Baker 1962: 222.
Diadromus contiguus, Chant & Yoshida-Shaul 1986: 2030, Moraes et al. 1986: 184.
Chanteius contiguus, Moraes et al. 2004: 261; Chant & McMurtry 1994: 239.
Chanteius lieni (Tseng 1976): 97 (synonymy according to Chant & Yoshida-Shaul 1986).
This species belongs to the contiguus species group (Chant & McMurtry 1994) and its biology remains totally unknown.
World distribution: China, Hong-Kong, Japan, Madagascar, Philippines, Singapore.
Specimens examined: 59 specimens in total, 51 ♀♀, 7 ♂♂ and 1 im. Coconi, Maison de L'Office National des Forêts (156 m aasl, 12°50'1'' S, 45°8'5'' E), 2 ♀♀ on Cinnamomum verum J. Presl (Lauraceae), 2 ♀♀ on Barleria lupulina Lindley (Acanthaceae), 2 ♀♀ on Stachytarpheta jamaicensis (L.) Vahl (Verbenaceae), 1 ♀ and 1 ♂ on Terminalia catappa L. (Combretaceae), 1 ♀ on Carica papaya L. (Caricaceae), 24/XI/2018; Miréréni (356 m aasl, 12°47'45'' S, 45°9'28'' E), 3 ♀♀ on Pteridium aqualinum (L.) Kuhn (Dennstaedtiaceae) and on Lantana camara L. (Verbenaceae), 24/XI/2018; Combani, gîte du Mont Combani (437 m aasl, 12°48'23'' S, 45°9'17'' E), 1 ♂ on Cananga odorata (Lamark) Hooker and Thomson (Annonaceae), 4 ♀♀, 1 ♂ and 1 im. on Citrus limon (L.) Burman (Rutaceae), 1 ♀ on C. papaya, 4 ♀♀ and 1 ♂ on Hydrangea aspera Buchanan-Hamilton ex D. Don (Hydrangeaceae), 2 ♀♀ on Annona muricata L. (Annonaceae), 14 ♀♀ and 1 ♂ on Cocos nucifera L. (Arecaceae), 7 ♀♀ on Bidens pilosa L. (Asteraceae), 1 ♀ and 1 ♂ on Artocarpus altilis (Parkinson) Fosberg (Moraceae) and 1 ♀ on Persea americana Miller (Lauraceae), 25/XI/2018; Coconi, Lycée Agricole (189 m aasl, 12°50'7'' S, 45°8'11'' E), 1 ♀ and 1 ♂ on C. papaya and 5 ♀♀ on Trema orientalis (L.) Blume (Cannabaceae), 26/XI/2018.
Remarks: morphological and morphometric characters and all measurements of our specimens fit well with measurements in numerous description and redescriptions available in the literature, especially those of Blommers (1976) for specimens from Madagascar. This is the most abundant species in our samplings. Mentioned only from South-East Asia and Madagascar, this is the first report of this species outside this main island.
Tribe Typhlodromini Wainstein
Typhlodromini Wainstein 1962: 26.
Genus Typhlodromus Scheuten
Typhlodromus Scheuten 1857: 111.
Subgenus Anthoseius De Leon
Typhlodromus (Anthoseius) De Leon 1959: 258.
Typhlodromus (Anthoseius) grewiae Zannou, Moraes & Oliveira
Typhlodromus (Anthoseius) grewiae Zannou, Moraes & Oliveira in Ueckermann et al. 2008: 48.
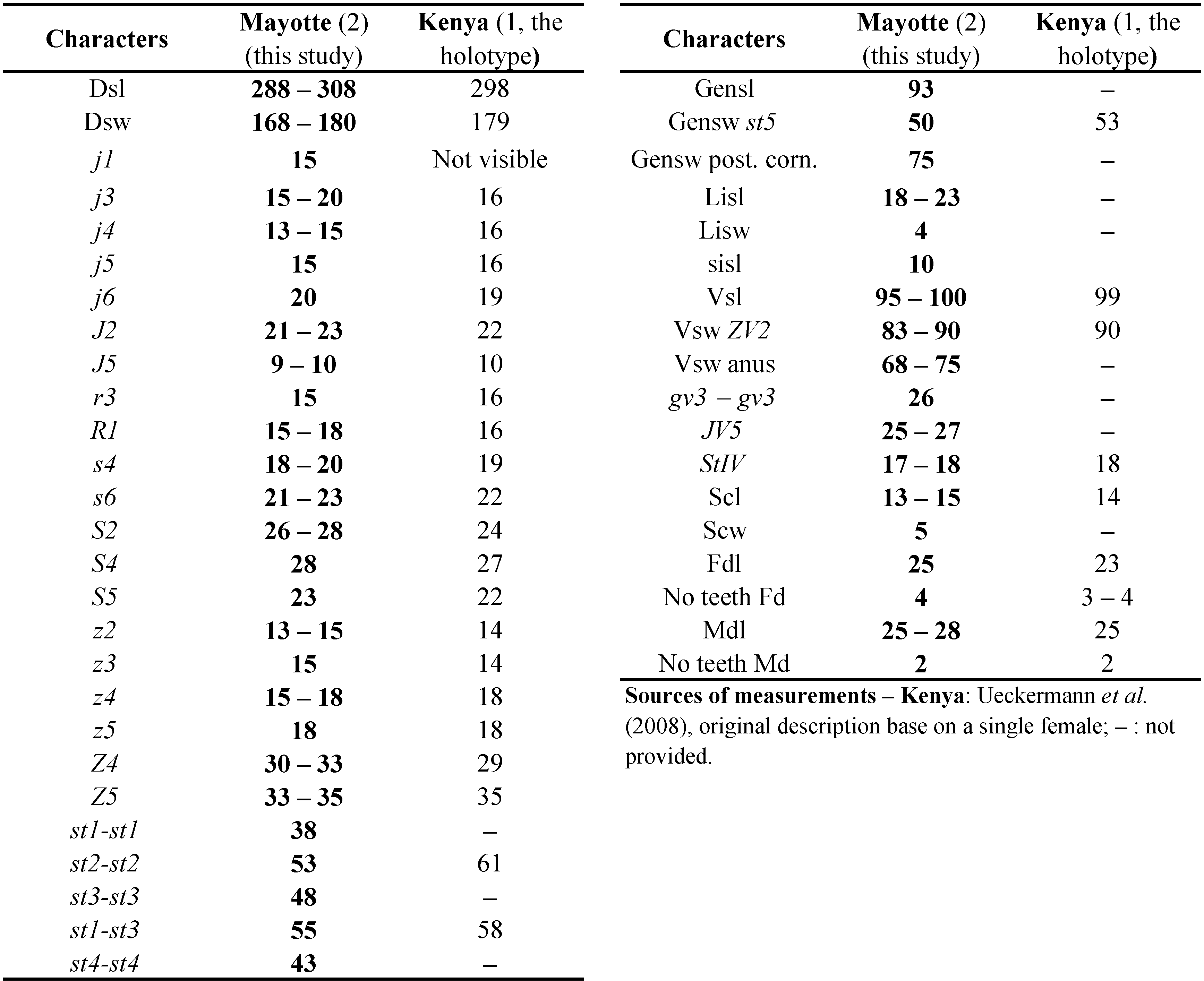
This species belongs to the singularis species group as setae JV3 are absent on the female and as specimens have shorter dorsal shield setae (Chant and McMurtry 1994). The biology of that species is totally unknown. It was reported only from Kenya (Ueckermann et al. 2008) based on a single female specimen.
World distribution: Kenya.
Specimens examined: 3 specimens in total, 2 ♀♀ and 1 ♂. Coconi, Maison de L'Office National des Forêts (156 m aasl, 12°50'1'' S, 45°8'5'' E), 2 ♀♀ on Terminalia catappa L. (Combretaceae), 24/XI/2018; Combani, gîte du Mont Combani (437 m aasl, 12°48'23'' S, 45°9'17'' E), 1 ♂ on Cananga odorata (Lamark) Hooker & Thomson (Annonaceae) or Ylang-Ylang, 25/XI/2018.
Remarks: the species was mentioned only once from Kenya and described based on a single female specimen. Morphological and morphometric characters and all measurements of our specimens (Table 6) fit well those of the original description by Ueckermann et al. (2008).
Typhlodromus (Anthoseius) lobatus Zannou, Moraes & Oliveira
Typhlodromus (Anthoseius) lobatus Zannou, Moraes & Oliveira in Ueckermann et al. 2008: 59.
This species belongs to the rhenanus species group (Chant and McMurtry 1994). The biology of that species is totally unknown. It was recently recorded from Rodrigues Island (Kreiter and Abo-Shnaf 2020a) and from Mauritius (Kreiter and Abo-Shnaf 2020b).
World distribution: Benin, Ghana, Mauritius Island, Rodrigues Island.
Specimens examined: 10 ♀♀ specimens in total. Coconi, Maison de L'Office National des Forêts (156 m aasl, 12°50'1'' S, 45°8'5'' E), 1 ♀ on Persea americana Miller (Lauraceae), 2 ♀♀ on Terminalia catappa L. Combretaceae) and 1 ♀ on an unknown host plant, 24/XI/2018; Combani, gîte du Mont Combani (437 m aasl, 12°48'23'' S, 45°9'17'' E), 1 ♀ on Bidens pilosa L. (Asteraceae), 25/XI/2018; Coconi, Lycée Agricole (189 m aasl, 12°50'7'' S, 45°8'11'' E), 1 ♀ on Capsicum annuum L. (Solanaceae), 2 ♀♀ on Rubus alceifolius Poiret (Rosaceae), 1 ♀ on Ageratum conizoides L. (Asteraceae), and 1 ♀ on Solanum mauritianum Scopoli (Solanaceae), 26/XI/2018.
Remarks: morphological and morphometric characters and all measurements of our specimens fit well with measurements of the original description by Ueckermann et al. (2008) concerning specimens from Ghana, in Western Africa and measurements of specimens from Rodrigues and Mauritius, respectively (Kreiter and Abo-Shnaf 2020a, b).
Typhlodromus (Anthoseius) microbullatus van der Merwe
Typhlodromus (Anthoseius) microbullatus van der Merwe 1968: 33; Moraes et al. 2004: 338; Chant & McMurtry 2007: 155; Ueckermann et al. 2008: 67.
Amblydromella microbullata, Moraes et al. 1986: 167.
Amblydromella (Aphanoseia) microbullata, Denmark & Welbourn 2002: 308.
This species belongs to the rhenanus species group (Chant and McMurtry 1994). The biology of that species is totally unknown. It was recorded from Madagascar, Mozambique and South Africa (Ueckermann et al. 2008).
World distribution: Madagascar, Mozambique, South Africa.
Specimens examined: 5 specimens in total, 3 ♀♀, 1 ♂ and 1 im. Combani, grower farm (104 m aasl, 12°47'14'' S, 45°7'57'' E), 1 ♀ on Theobroma cacao L. (Malvaceae), 26/XI/2018; L'Abattoir, Dziani lake (23 m aasl, 12°46'14'' S, 45°17'18'' E), 1 ♀, 1 ♂ and 1 im. on Artocarpus altilis (Parkinson) Fosberg (Moraceae) and 1 ♀ on Tamarindus indica L. (Fabaceae), 27/XI/2018.
Remarks: morphological and morphometric characters and all measurements of our specimens fit well those of specimens from South Africa in van der Merwe (1968) and Ueckermann et al. (2008).
Conclusion
The results of an additional survey made in 2018 in Mayotte Island is presented in this paper. A total of 18 new records: 13 Amblyseiinae, 1 Phytoseiinae and 4 Typhlodrominae, have been obtained, namely Neoseiulus barkeri, N. teke, Paraphytoseius horrifer, P. orientalis, Typhlodromips shi, Amblyseius largoensis, A. parasundi, A. tamatavensis, Proprioseiopsis ovatus, Euseius ovaloides, Moraeseius papayana, Phytoseius haroldi, Chanteius contiguus, Typhlodromus (Anthoseius) lobatus, T. (A.) grewiae, T. (A.) microbullatus. Two new species will be described in a following paper.
The fauna of Mayotte after our study is composed of 19 species: 13 Amblyseiinae, 2 Phytoseiinae and 4 Typhlodrominae. Unfortunately, we have not recovered the unique species known before our study, Phytoseius mayottae Schicha. Among the 18 newly recorded species, at least four species (N. barkeri, A. largoensis, A. tamatavensis, and E. ovaloides) are known as biological control agents (BCAs). In addition to the intrinsic value of phytoseiid mite biodiversity in tropical environments, demonstration of the natural occurrence of efficient BCAs in a developing country such as Mauritius is of great agricultural, commercial and strategic interests for the country.
Acknowledgements
Thanks are first due to the Department to which the senior author belong for research that have granted travels and accommodations to him in Mauritius: UMR CBGP (Internal call for proposals 2018). A grateful thanks to UR Hortsys and to the head Dr Fabrice Le Bellec that have allowed Rose-My Payet to join the senior author for collections and partly granted field collecting trips. Thanks also to the I-SITE Montpellier Université d'Excellence (MUSE) for the international mobility support to Reham I.A. Abo-Shnaf (Explore #2, the MUSE International Mobility program 2019). Thanks finally to Karl-Heinz Schmidt for information about taxonomic data concerning the species Paraphytoseius orientalis.
References
Athias-Henriot C. 1960. Phytoseiidae et Aceosejidae (Acarina: Gamasina) d$'$Algérie. IV. Genre Typhlodromus Scheuten, 1857. Bull. Soc. Hist. Nat. Afr. Nord, 51: 62-107.
Athias-Henriot C. 1961. Mésostigmates (Urop. excl.) édaphiques Méditerranéens (Acaromorpha, Anactinotrichida). Acarologia, 3: 381-509.
Athias-Henriot C. 1975. Nouvelles notes sur les Amblyseiini. II. Le relevé organotaxique de la face dorsale adulte (Gamasides protoadéniques, Phytoseiidae). Acarologia, 17: 20-29.
Beard J.J. 2001. A review of Australian Neoseiulus Hughes and Typhlodromips De Leon (Acari: Phytoseiidae: Amblyseiinae). Invert. Taxon., 15: 73-158. doi:10.1071/IT99017 ![]()
Beaulieu F., Beard J.J. 2018. Acarine biocontrol agents Neoseiulus californicus sensu Athias-Henriot (1977) and N. barkeri Hughes redescribed, their synonymies assessed, and the identity of N. californicus clarified based on examination types. Zootaxa, 4500: 451-507. doi:10.11646/zootaxa.4500.4.1 ![]()
Berlese A. 1913. Systema Acarorum genera in familiis suis disposita. Acaroteca Italica, 1-2: 3-19.
Berlese A. 1914. Acari nuovi. Manipulus IX. Redia, 10: 113-150.
Blommers L. 1973. Five new species of phytoseiid mites (Acarina: Phytoseiidae) from southwest Madagascar. Bull. Zool. Mus. Univ. Amst., 3: 109-117.
Blommers L. 1974. Species of the genus Amblyseius Berlese, 1914, from Tamatave, east Madagascar (Acarina: Phytoseiidae). Bull. Zool. Mus. Univ. Amst., 3: 143-155.
Blommers L. 1976. Some Phytoseiidae (Acarina: Mesostigmata) from Madagascar, with descriptions of eight new species and notes on their biology. Bijdragen tot Dierkunde, 46: 80-106. doi:10.1163/26660644-04601005 ![]()
Blommers L., Gutierrez J. 1975. Les tétranyques vivant sur agrumes et avocatiers dans la région de Tamatave (Madagascar - est) et quelques-uns de leurs prédateurs. Fruits, 30: 191-200.
Broodsgaard H.F., Stengaard Hansen L. 1992. Effect of Amblyseius cucumeris and Amblyseius barkeri as Biological Control Agents of Thrips tabaci on Glasshouse Cucumbers. Biocont. Sci. Technol., 2: 215-223. doi:10.1080/09583159209355235 ![]()
Blommers L., Chazeau J. 1974. Two new species of predator mites of the genus Amblyseius Berlese (Acarina: Phytoseiidae) from Madagascar. Zeit. Ang. Entomol., 75: 308-315. doi:10.1111/j.1439-0418.1974.tb01856.x ![]()
Cavalcante, A.C.C., Mandro, M.E.A., Romanini, E.P., Moraes, G.J. de. 2017. Amblyseius tamatavensis Blommers (Acari: Phytoseiidae) a candidate for biological control of Bemisia tabasi (Gennadius) biotype B (Hemiptera: Aleyrodidae) in Brazil. Int. J. Acarol., 43: 10-15. doi:10.1080/01647954.2016.1225816 ![]()
Chant D.A. 1959. Phytoseiid mites (Acarina: Phytoseiidae). Part I. Bionomics of seven species in southeastern England. Part II. A taxonomic review of the family Phytoseiidae, with descriptions of thirty-eight new species. Can. Entomol., 61: 1-166. doi:10.4039/entm9112fv ![]()
Chant D.A., Hansell R.I.C. 1971. The genus Amblyseius in Canada and Alaska. Can. J. Zool., 49: 703-758. doi:10.1139/z71-110 ![]()
Chant D.A., McMurtry J.A. 1994. A review of the subfamilies Phytoseiinae and Typhlodrominae (Acari: Phytoseiidae). Int. J. Acarol., 20: 223-310. doi:10.1080/01647959408684022 ![]()
Chant D.A., McMurtry J.A. 2003a. A review of the subfamily Amblyseiinae Muma (Acari: Phytoseiidae): Part I. Neoseiulini new tribe. Int. J. Acarol., 29: 3-46. doi:10.1080/01647950308684319 ![]()
Chant D.A., McMurtry J.A. 2003b. A review of the subfamily Amblyseiinae Muma: Part II. The tribe Kampimodromini Kolodochka. Int. J. Acarol., 29: 179-224. doi:10.1080/01647950308684331 ![]()
Chant D.A., McMurtry J.A. 2004. A review of the subfamily Amblyseiinae Muma (Acari: Phytoseiidae): Part III. The tribe Amblyseiini Wainstein, subtribe Amblyseiina n. subtribe. Int. J. Acarol., 30: 171-228. doi:10.1080/01647950408684388 ![]()
Chant D.A., McMurtry J.A. 2005a. A review of the subfamily Amblyseiina Muma (Acari: Phytoseiidae): Part V. Tribe Amblyseiini, subtribe Proprioseiopsina Chant and McMurtry. Int. J. Acarol., 31: 3-22. doi:10.1080/01647950508684412 ![]()
Chant D.A., McMurtry J.A. 2005b. A review of the subfamily Amblyseiinae Muma (Acari: Phytoseiidae) Part VI. The tribe Euseiini n. tribe, subtribes Typhlodromalina n. subtribe, Euseiina n. subtribe, and Ricoseiina n. subtribe. Int. J. Acarol., 31: 187-224. doi:10.1080/01647950508684424 ![]()
Chant D.A., McMurtry J.A. 2005c. A review of the subfamily Amblyseiinae Muma (Acari: Phytoseiidae) Part VII. Typhlodromipsini n. tribe. Int. J. Acarol., 31: 315-340. doi:10.1080/01647950508683673 ![]()
Chant D.A., McMurtry J.A. 2006. A review of the subfamily Amblyseiinae Muma (Acari: Phytoseiidae): Part IX. An overview. Int. J. Acarol., 32: 1-27. doi:10.1080/01647950608684439 ![]()
Chant D.A., McMurtry J.A. 2007. Illustrated keys and diagnoses for the genera and subgenera of the Phytoseiidae of the world (Acari: Mesostigmata). Indira Publishing House, West Bloomfield, 219 pp.
Chant D.A., Yoshida-Shaul E. 1986. A world review of the ecclesiasticus species group in the genus Typhlodromus Scheuten (Acarina: Phytoseiidae). Can. J. Zool., 64: 447-466. doi:10.1139/z86-069 ![]()
Chant D.A., Yoshida-Shaul E. 1991. Adult ventral setal patterns in the family Phytoseiidae (Acari: Gamasina). Int. J. Acarol., 17: 187-199. doi:10.1080/01647959108683906 ![]()
Chant D.A., Yoshida-Shaul E. 1992. Adult idiosomal setal patterns in the family Phytoseiidae (Acari: Gamasina). Int. J. Acarol., 18: 177-193. doi:10.1080/01647959208683949 ![]()
Congdon B.D. 2002. The family Phytoseiidae (Acari) in western Washington State with descriptions of three new species. Int. J. Acarol., 28: 3-27. doi:10.1080/01647950208684275 ![]()
De Leon D. 1959. Two new genera of phytoseiid mites with a note on Proprioseius meridionalis Chant (Acarina: Phytoseiidae). Entomol. News, 70: 257-262.
De Leon D. 1966. Phytoseiidae of British Guyana with keys to species (Acarina: Mesostigmata). Stud. Fauna Suriname and other Guyanas, 8: 81-102.
Demite P.R., McMurtry J.A., Moraes G.J. de. 2014. Phytoseiidae Database: a website for taxonomic and distributional information on phytoseiid mites (Acari). Zootaxa, 3795: 571-577. doi:10.11646/zootaxa.3795.5.6 ![]()
Demite P.R., Moraes G.J. de, McMurtry J.A., Denmark H.A., Castilho R.C. 2020. Phytoseiidae Database. Available from: www.lea.esalq.usp.br/phytoseiidae ![]() (last access 30/05/2020).
(last access 30/05/2020).
Denmark H.A., Evans G.A. 2011. Phytoseiidae of North America and Hawaii (Acari: Mesostigmata). Indira Publishing House, West Bloomfield, USA, 451 pp.
Denmark H.A., Muma M.H. 1989. A revision of the genus Amblyseius Berlese, 1914 (Acari: Phytoseiidae). Occas. Pap. Fla State Coll. Arthropods, USA, 4, 149 pp.
Denmark H.A., Welbourn W.C. 2002. Revision of the genera Amblydromella Muma and Anthoseius De Leon (Acari: Phytoseiidae). Int. J. Acarol., 28: 291-316. doi:10.1080/01647950208684308 ![]()
Denmark H.A., Evans G.A., Aguilar H., Vargas C., Ochoa R. 1999. Phytoseiidae of Central America (Acari: Mesostigmata). Indira Publishing House, West Bloomfield, Michigan, USA. 125 pp.
Ehara S. 1966. A tentative catalogue of predatory mites of Phytoseiidae known from Asia, with descriptions of five new species from Japan. Mushi, 39: 9-30.
Ehara S. 1967. Phytoseiid mites from Okinawa Island (Acarina: Mesostigmata). Mushi, 40: 67-82.
Ehara S. 2002. Some phytoseiid mites (Arachnida: Acari: Phytoseiidae) from west Malaysia. Species Div., 7: 29-46. doi.org/10.12782/specdiv.7.29 doi:10.12782/specdiv.7.29 ![]()
Ehara S., Amano H. 2002. Some Japanese phytoseiid mites (Acari: Phytoseiidae) mostly from Ishigaki and Taketomi Islands. Entomol. Sci., 5: 321-329.
Ehara S., Amano H. 2004. Checklist and keys to Japanese Amblyseiinae (Acari: Gamasina: Phytoseiidae). J. Acarol. Soc. Japan, 13: 1-30. doi:10.2300/acari.13.1 ![]()
El-Banhawy E.M. 1984. Description of some phytoseiid mites from Brazil. Acarologia, 25: 125-144.
El-Banhawy E.M., Knapp M. 2011. Mites of the family Phytoseiidae Berlese from Kenya (Acari: Mesostigmata). Zootaxa, 2945: 1-176. doi:10.11646/zootaxa.2945.1.1 ![]()
Fan Y.Q., Petitt F.L. 1994a. Biological Control of Broad Mite, Polyphagotarsonemus latus (Banks), by Neoseiulus barkeri Hughes on Pepper. Biol. Cont., 4: 390-395. doi:10.1006/bcon.1994.1049 ![]()
Fan Y.Q., Petitt F.L. 1994b. Parameter estimation of the functional response. Environ. Entomol., 23: 785-794. doi:10.1093/ee/23.4.785 ![]()
Ferragut F., Baumann J. 2019. New phytoeiid mites (Mesostigmata: Phytoseiidae) of Mauritius, with the description of two new species. Syst. Appl. Acarol., 24: 825-856. doi:10.11158/saa.24.5.8 ![]()
Furtado I.P., Moraes G.J. de, Kreiter S., Flechtmann C.H.W., Tixier M.-S., Knapp M. 2014. Plant inhabiting phytoseiid predators of Midwestern Brazil, with emphasis on those associated with the tomato red spider mite, Tetranychus evansi. Acarologia, 54: 425-431. doi:10.1051/acarologia/20142138 ![]()
Garman P. 1958. New species belonging to the genera Amblyseius and Amblyseiopsis with keys to Amblyseius, Amblyseiopsis, and Phytoseiulus. Ann. Entomol. Soc. Amer., 51: 69-79. doi:10.1093/aesa/51.1.69 ![]()
Gupta S.K. 1977. Some undescribed and little-known species of Amblyseius (Acarina: Phytoseiidae) from western and northern India. Ind. J. Acarol., 1: 28-37.
Gupta S.K. 1986. Fauna of India. Family Phytoseiidae. Zoological Survey of India, Calcutta, India, 350 pp.
Gutierrez J., Etienne J. 1986. Les Tetranychidae de I$'$île de la Réunion et quelques-uns de leurs prédateurs. Agron. Trop., 41: 84-91.
Hirschmann W. 1962. Gangystematik der Parasitiformes. Acarologie Schriftenreihe fur Vergleichende Milbenkunde, Hirschmann-Verlag, Furth/Bay, 5: 80 pp.+ 32 plates.
Hughes A.M. 1948. The mites associated with stored food products. Ministry of Agriculture and Fisheries, H. M. Stationary Office, London, 168 pp.
Hughes A.M. 1961. The mites of stored food. Ministry of Agriculture, Fishery and Food Technical Bulletin, First Edition, London, 9: 1-287.
Karg W. 1970. Neue Arten der Raubmilbenfamilie Phytoseiidae Berlese, 1916 (Acarina: Parasitiformes). Deut. Entomol. Zeit. N. F., 17: 289-301. doi.org/10.1002/mmnd.4810170402 doi:10.1002/mmnd.4810170402 ![]()
Karg W. 1982. Diagnostic and systematics of predatory mites of the family Phytoseiidae Berlese in orchards. Zool. Jahrb. Syst., 109: 188-210.
Karg W. 1983. Systematische untersuchung der Gattungen und Untergattungen der Raubmilbenfamilie Phytoseiidae Berlese, 1916, mit der beschreibung von 8 neuen Arten. Mitt. Zool. Mus. Berlin, 59: 293-328. doi.org/10.1002/mmnz.4830590203 doi:10.1002/mmnz.4830590203 ![]()
Karg W. 1989. Neue Raubmilbenarten der Gattuig Proprioseiopsis Muma, 1961 (Acarina, Parasitiformes) mit Bestimmungsschlüsseln. Zool. Jahrb. Syst., 116: 199-216.
Kolodochka L.A. 1998. Two new tribes and the main results of a revision of Paleartic phytoseiid mites (Parasitiformes, Phytoseiidae) with the family system concept. Vest. Zool., 32: 51-63 [in Russian].
Kreiter S., Abo-Shnaf R.I.A. 2020a. Phytoseiid mites of Rodrigues Island. Acarologia, 60: 449-468. doi:10.24349/acarologia/20204376 ![]()
Kreiter S., Abo-Shnaf R.I.A. 2020b. New records of phytoseiid mites from Mauritius Island (Acari: Mesostigmata). Acarologia, 60: 520-545. doi:10.24349/acarologia/20204382 ![]()
Kreiter S., Amiri K., Douin M., Bohinc T., Trdan S., Tixier M.-S. 2020a. Phytoseiid mites of Slovenia (Acari: Mesostigmata): new records and first description of the male of Amblyseius microorientalis. Acarologia, 60: 203-242. doi:10.24349/acarologia/20204364 ![]()
Kreiter S., Bopp M.-C., Douin M., Nguyen D.T., Wyckhuys K. 2020b. Phytoseiidae of Vietnam with description of a new species. Acarologia, 60: 75-110. doi:10.24349/acarologia/20204362 ![]()
Kreiter S., Fontaine O., Payet R.-M. 2018a. New records of Phytoseiidae (Acari: Mesostigmata) from Mauritius. Acarologia, 58: 773-785. doi:10.24349/acarologia/20184273 ![]()
Kreiter S., Moraes G.J. de. 1997. Phytoseiidae mites (Acari: Phytoseiidae) from Guadeloupe and Martinique. Fla Entomol., 80: 376-382. doi:10.2307/3495770 ![]()
Kreiter S., Payet R.-M., Douin M., Fontaine O., Fillâtre J., Le Bellec F. 2020c. Phytoseiidae of La Réunion Island (Acari: Mesostigmata): three new species and two males described, new synonymies, and new records. Acarologia, 60: 111-195. doi:10.24349/acarologia/20204361 ![]()
Kreiter S., Payet R.-M., Fillâtre J., Abdou Azali H. 2018b. First records of Phytoseiidae from one island of the Comoros Archipelago. Acarologia, 58: 529-545. doi:10.24349/acarologia/20184256 ![]()
Kreiter S., Tixier M.-S., Etienne J. 2006. New records of phytoseiid mites from the French Antilles, with description of Neoseiulus cecileae sp. nov. Zootaxa, 1294: 1-27. doi:10.11646/zootaxa.1294.1.1 ![]()
Kreiter S., Ueckermann E.A., Quilici S. 2002. Seven new phytoseiid species, with a new generic assignement and a key to the species of La Reunion Island (Acari: Mesostigmata). Acarologia, 42: 335-350.
Kreiter S., Zriki Z., Ryckewaert P., Pancarte C., Douin M., Tixier M.-S. 2018c. New phytoseiid mites of Martinique, with redescription of four species and new records. Acarologia, 58: 366-407. doi:10.24349/acarologia/20184248 ![]()
Lindquist E.E. 1994. Some observations on the chaetotaxy of the caudal body region of gamasine mites (Acari: Mesostigmata), with a modified notation for some ventrolateral body setae. Acarologia, 35: 323-326.
Lindquist E.E., Evans G.W. 1965. Taxonomic concepts in the Ascidae, with a modified setal nomenclature for the idiosoma of the Gamasina. Mem. Entomol. Soc. Canada, 47: 1-64. doi:10.4039/entm9747fv ![]()
Mailloux J., Le Bellec F., Kreiter S., Tixier M.-S., Dubois P. 2010. Influence of ground cover management on diversity and density of phytoseiid mites (Acari: Phytoseiidae) in Guadeloupean citrus orchards. Exp. Appl. Acarol., 52: 275-290. doi:10.1007/s10493-010-9367-7 ![]()
Massaro M., Montrazi M., Melo J.W.S., Moraes G.J. 2018. Production of Amblyseius tamatavensis with Thyreophagus crasentiseta (Acari: Phytoseiidae, Acaridae). Int. J. Pest Manag. (in press).
Massaro M. Moraes G.J. de. 2019. Predation and oviposition potential of Brazilian populations of the predatory mite Amblyseius tamatavensis (Acari: Phytoseiidae) on eggs of Bemisia tabaci (Insecta: Hemiptera). Acarologia, 59: 120-128. doi:10.24349/acarologia/20194314 ![]()
Matthysse J.G., Denmark H.A. 1981. Some phytoseiids of Nigeria (Acarina: Mesostigmata). Fla Entomol., 64: 340-357. doi:10.2307/3494585 ![]()
McMurtry J.A., Croft B.A. 1997. Life-styles of phytoseiid mites and their roles in biological control. Ann. Rev. Entomol., 42: 291-321. doi:10.1146/annurev.ento.42.1.291 ![]()
McMurtry J.A., Moraes G.J. de, Sourassou N.F. 2013. Revision of the life styles of phytoseiid mites (Acari: Phytoseiidae) and implications for biological control strategies. Syst. Appl. Acarol., 18: 297-320. doi:10.11158/saa.18.4.1 ![]()
Meyer M.K.P., Rodrigues M. da C. 1966. Acari associated with Cotton in Southern Africa. References to other plants. Garcia de Orta, Rev. Junta Investig., 13: 27-31.
Moraes G.J. de, Castro T.M.G. de, Kreiter S., Quilici S., Gondim Jr. M.G.C., Sá L.A. 2012. Search for natural enemies of Raoiella indica Hirst in Réunion Island. Acarologia, 52: 129-134. doi:10.1051/acarologia/20122043 ![]()
Moraes G.J. de, Kreiter S., Lofego A.C. 2000. Plant mites (Acari) of the French Antilles. 3. Phytoseiidae (Gamasida). Acarologia, 40: 237-264.
Moraes G.J. de, McMurtry J.A., Denmark H.A. 1986. A catalog of the mite family Phytoseiidae. References to taxonomy, synonymy, distribution and habitat. EMBRAPA - DDT, Brasilia, Brazil, 353 pp.
Moraes G.J. de, McMurtry J.A., Denmark H.A., Campos C.B. 2004. A revised catalog of the mite family Phytoseiidae. Zootaxa, 434: 1-494. doi:10.11646/zootaxa.434.1.1 ![]()
Moraes G.J. de, McMurtry J.A., van den Berg H., Yaninek J.S. 1989a. Phytoseiid mites (Acari: Phytoseiidae) of Kenya, with descriptions of five new species and complementary descriptions of eight species. Int. J. Acarol., 15: 79-93. doi:10.1080/01647958908683829 ![]()
Moraes G.J. de, McMurtry J.A., Yaninek, J.S. 1989b. Some phytoseiid mites from tropical Africa with description of a new species. Int. J. Acarol., 15: 95-102. doi:10.1080/01647958908683830 ![]()
Moraes G.J. de, Ueckermann E.A., Oliveira A.R., Yaninek J.S. 2001. Phytoseiidae mites of the genus Euseius (Acari: Phytoseiidae) from Sub-Saharan Africa. Zootaxa, 3: 1-70. doi:10.11646/zootaxa.3.1.1 ![]()
Moraes G.J. de, Zannou I.D., Ueckermann E.A., Oliveira A.R., Hanna R., Yaninek J.S. 2007. Species of the subtribes Arrenoseiina and Proprioseiopsina (Tribe Amblyseiini) and the tribe Typhlodromipsini (Acari: Phytoseiidae) from sub-Saharan Africa. Zootaxa, 1448: 1-39. doi:10.11646/zootaxa.1448.1.1 ![]()
Muma M.H. 1955. Phytoseiidae associated with citrus in Florida. Ann. Entomol. Soc. Amer., 48: 262-272. doi:10.1093/aesa/48.4.262 ![]()
Muma M.H. 1961. Subfamiles, genera, and species of Phytoseiidae. Fla St. Mus. Bul., 5: 267-302.
Muma M.H. 1962. New Phytoseiidae (Acarina: Mesostigmata) from Florida. Florida Entomol., 45: 1-10. doi:10.2307/3492897 ![]()
Myers N. 1988. Threatened biotas: hostspots in tropical forests. Environmentalist, 8: 187-208. doi.org/10.1007/BF02240252 doi:10.1007/BF02240252 ![]()
Myers N., Mittermeier R.A., Mittermeier C.G., Da Fonseca G.A., Kent J. 2000. Biodiversity hotspots for conservation priorities. Nature, 403: 853-858. doi:10.1038/35002501 ![]()
Narayanan E.S., Kaur R.B.N., Ghai S. 1960. Importance of some taxonomic characters in the family Phytoseiidae., with new records and descriptions of species. Proceed. Nat. Inst. Sc. India, 26B: 384-394.
Navasero M.M., Navasero M.V. 2016. Biology of Paraphytoseius orientalis (Narayanan et al.) reared on the broad mite, Polyphagotarsonemus latus (Banks) in the Philippines. Philip. Entomol., 30: 21-28.
Navia D., Domingos C.A., Mendonça R.S., Ferragut F., Rodrigues M.A.N., de Morais E.G.F., Tixier M.-S., Gondim Jr. M.G.C. 2014. Reproductive compatibility and genetic and morphometric variability among populations of the predatory mite, Amblyseius largoensis (Acari: Phytoseiidae), from Indian Ocean Islands and the Americas. Biol. Cont., 72: 17-29. doi:10.1016/j.biocontrol.2014.01.011 ![]()
Nesbitt H.H.J. 1951. A taxonomic study of the Phytoseiinae (Family Laelaptidae) predaceous upon Tetranychidae of economic importance. Zool. Verhandel., 12: 1-96.
Nwilene F.E., Nachman G. 1996. Functional responses of Iphiseius degenerans and Neoseiulus teke (Acari: Phytoseiidae) to changes in the density of the cassava green mite, Mononychellus tanajoa (Acari: Tetranychidae). Exp. Appl. Acarol., 20: 259-271. doi:10.1007/BF00052876 ![]()
Prasad V. 1968. Amblyseius mites from Hawaii. Ann. Entomol. Soc. Amer., 61: 1514-1521. doi:10.1093/aesa/61.6.1514 ![]()
Pritchard A.E., Baker E.W. 1962. Mites of the family Phytoseiidae from Central Africa, with remarks on the genera of the world. Hilgardia, 33: 205-309. doi:10.3733/hilg.v33n07p205 ![]()
Quilici S., Kreiter S., Ueckermann E. A., Vincenot D. 1997. Predatory mites (Acari) from various crops on Réunion Island. Int. J. Acarol., 23: 283-291. doi:10.1080/01647959708683578 ![]()
Quilici S., Ueckermann E. A., Kreiter S., Vayssières J.-F. 2000. Phytoseiidae (Acari) of La Réunion Island. Acarologia, 41: 97-108.
Ragusa S., Athias-Henriot C. 1983. Observations on the genus Neoseiulus Hughes (Parasitiformes, Phytoseiidae). Redefinition. Composition. Geography. Description of two new species. Rev. Suisse Zool. 90: 657-678. doi:10.5962/bhl.part.82005 ![]()
Ribaga C. 1904. Gamasidi planticoli. Riv. Patol. Veget., 10: 175-178.
Rodriguez-Reina J.M., Garcia-Mari F., Ferragut F. 1992. Predatory activity of phytoseiid mites on different developmental stages of the Western flower thrips Frankliniella occidentalis. Bol. Sanid. Veget. Plagas, 18: 253-263.
Rowell H.J., Chant D.A., Hansell R.I.C. 1978. The determination of setal homologies and setal patterns on the dorsal shield in the family Phytoseiidae. Can. Entomol., 110: 859-876. doi:10.4039/Ent110859-8 ![]()
Ryu M.O., Lee W.K., Cho S.R. 2001. Phytoseiid mites (Acari: Phytoseiidae) from the pear field of Naju District in Korea. Kor. J. Soil Zool., 6: 7-9.
Scheuten A. 1857. Einiges uber Milben. Archiv für Naturgeschichte, 23: 104-112.
Schicha E. 1984. Contribution to the knowledge of the genus Phytoseius in Australia, the South Pacific and Indian Ocean regions with four new species and records of known species. Int. J. Acarol., 10: 117-128. doi:10.1080/01647958408683361 ![]()
Schicha E. 1987. Phytoseiidae of Australia and neighbouring areas. Indira Publishing House, West Bloomfield, Michigan, USA, 187 pp.
Schicha E., Gutierrez J. 1985. Phytoseiidae of Papua New Guinea, with three new species, and new records of Tetranychidae (Acari). Int. J. Acarol., 11: 173-181. doi:10.1080/01647958508683412 ![]()
Schuster R.O., Pritchard A.E. 1963. Phytoseiid mites of California. Hilgardia, 34: 191-285. doi:10.3733/hilg.v34n07p191 ![]()
Specht H.B. 1968. Phytoseiidaein the New Jersey apple orchard environment with descriptions of spermathecae and three new species. Can. Entomol., 100: 673-692. doi:10.4039/Ent100673-7 ![]()
Swirski E., Golan Y. 1967. On some phytoseiid mites (Acarina) from Luzon Island (Philippines). Israel J. Agric. Res., 17: 225-227.
Swirski E., Shechter R. 1961. Some phytoseiid mites (Acarina: Phytoseiidae) of Hong-Kong, with a description of a new genus and seven new species. Isr. J. Agric. Res., 11: 97-117.
Tseng Y.H. 1976. Systematics of the mite family Phytoseiidae from Taiwan, with a revised key to genera of the world (II). J. Agric. Ass. China New Series, 94: 85-128.
Tseng Y.H. 1983. Further study on phytoseiid mites from Taiwan. Chin. J. Entomol., 3: 33-74.
Ueckermann E.A., Loots G.C. 1988. The African species of the subgenera Anthoseius De Leon and Amblyseius Berlese (Acari: Phytoseiidae). Entomol. Mem., Dep. Agric. Water Supply, Rep. South Africa, 73, 168 pp.
Ueckermann, E.A., Zannou, I.D., Moraes, G.J. de, Oliveira, A.R. de, Hanna, R., Yaninek J.S. 2008. Phytoseiid mites of the tribe Typhlodromini (Acari: Phytoseiidae) from sub-Saharan Africa. Zootaxa, 1901: 1-122. doi:10.11646/zootaxa.1901.1.1 ![]()
van der Merwe G.G. 1965. South African Phytoseiidae (Acarina). I. Nine new species of the genus Amblyseius Berlese. J. Entomol. Soc. South Afr., 28: 57-76.
van der Merwe G.G. 1968. A taxonomic study of the family Phytoseiidae (Acari) in South Africa with contributions to the biology of two species. Entomol. Mem. South Africa Dep. Agric. Techn. Serv., 18: 1-198.
Vitzthum H. von 1941. Acarina. In: Bronns, H.G. (Ed.), Klassen und Ordnungen des Tierreichs 5, Akademischer Verlag, Leipzig, Germany, pp. 764-767.
Wainstein B.A. 1962. Révision du genre Typhlodromus Scheuten, 1857 et systématique de la famille des Phytoseiidae (Berlese 1916) (Acarina: Parasitiformes). Acarologia, 4: 5-30.
Wu, W.N., Chou, F.W. 1981. A new species of Amblyseius (Acarina: Phytoseiidae) from Guangdong Province [in Chinese]. Zoological Research, 2: 273-274.
Zannou I.D., Moraes G.J. de, Ueckermann E.A., Oliveira A.R., Yaninek J.S., Hanna R. 2006. Phytoseiid mites of the genus Neoseiulus Hughes (Acari: Phytoseiidae) from sub-Saharan Africa. Int. J. Acarol., 32: 241-276. doi:10.1080/01647950608684467 ![]()
Zannou I.D., Moraes G.J. de, Ueckermann E.A., Oliveira A.R., Yaninek J.S., Hanna R. 2007. Phytoseiid mites of the subtribe Amblyseiina from sub-Saharan Africa. Zootaxa, 1550: 1-47. doi:10.11646/zootaxa.1550.1.1 ![]()



2020-07-07
Date accepted:
2020-08-18
Date published:
2020-09-08
Edited by:
Faraji, Farid

This work is licensed under a Creative Commons Attribution 4.0 International License
2020 Kreiter, Serge; Abo-Shnaf, Reham I.A. and Payet, Rose-My
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)