



Occigamasus, a new genus of pergamasine mites, with description of two new species from the west coast of North America (Parasitiformes: Gamasina: Parasitidae)
Juvara-Balş, Ilinca1
1Muséum d’histoire naturelle, CP 6434, Genève 6. Switzerland.
2019 - Volume: 59 Issue: 4 pages: 551-570
https://doi.org/10.24349/acarologia/20194354ZooBank LSID: D9D1B3C7-98C9-4284-9F53-8D451D4746E5
Original research
Keywords
Abstract
Introduction
Predatory mites of the Family Parasitidae Oudemans, 1901 are important soil and litter inhabitants in the Holarctic region. The family was divided into two subfamilies Parasitinae Oudemans, 1901 and Pergamasinae Juvara-Balş, 1972. Various studies of parasitid mites have been done in Europe (Berlese 1906; Bhattacharyya 1963; Athias-Henriot 1967a, 1967b, 1967c, 1971; Micherdzinsky 1969, Hyatt 1980). However, the diversity of these mites in North America has been scarcely investigated with the most important works being those of Banks (1904, 1910) and Ewing (1909).
Hennessey and Farrier (1988, 1989) re-examined and further described and illustrated the species of Parasitidae described by Banks and Ewing. Initially a systematic revision of thirty species in ten families of gamasine mites was published (1988), including seven species in six genera of Parasitidae, followed by a paper concerning only the family Parasitidae (1989) from North and South Carolina. In the latter paper, fourteen species, including one new genus and four new species were described, and new synonymies, new combination and a lectotype designated.
Scant information is available about parasitid mites in Canada. Richards (1976) published a paper concerning Parasitidae mites associated with bumblebees in Alberta. An estimated 35 species representing 6 genera of Parasitidae were recorded, without attributed names, as occurring in Canada (Lindquist et al. 1979); and that number of species was increased to 51, with almost twice as many species estimated to await discovery (Beaulieu et al. 2019). This paper deals with Pergamasinae mites from North America: Vancouver Island, British Columbia and Alberta (Canada) as well as Oregon (USA). Here, I describe an unusual new genus, Occigamasus and two new species, O. lindquisti and O. makarovae. The sites of other undescribed species belonging to the new genus, which need more specimens to be described adequately, are indicated.
Material and Methods
The mites studied (52 slides) are from the Canadian National Collection (CNC) deposited in the Research and Development center, Agriculture and Agri-Food, Ottawa. E. E. Lindquist collected most of the mites from Vancouver Island and Oregon. L. Russel collected one sample from Oregon and A. Fjellberg another one from Comox Glacier – Vancouver Island.
The specimens were mounted in Hoyer's medium and became flattened, so that not all the characters are easily visible. The morphological terminology and the system of setal notation for the chelicerae and palpi follows Evans and Till (1979). Terminology for the leg chaetotaxy is based on Evans (1963). The setal notation for the idiosomal dorsum follows Lindquist and Evans (1965) and that for the opisthogaster follows Lindquist (1994). The notation for adenotaxy is based on the system presented by Johnston and Moraza (1991) which is slightly different from that established by Athias-Henriot (1969a). For comparison with other papers the nomenclature of Athias-Henriot (1969a) is given in parentheses: gdj2 (gd1); gdj4 (gd2); gds3 (gd3); gds4 (gd4); gdz5 (gd5); gdz6 (gd6); gdZ2 (gd7); gdZ3 (gd8); gdZ4 (gd9). Measurements of female structures: epigynium shield height (h) is the length from the tip of the shield to its posterior margin, epigynium basal width (b) represents the length of the posterior margin; setal distance st–st is a transverse measurement between insertions of the two setae of the pair st on sternal shield. Leg length is taken from the base of the coxa to the apex of the tarsus; tarsus I and IV length were taken from the base of the segment to its apex, without the pretarsus. Measurements in micrometers are specified after each structure. The following abbreviations are used: hyp. hypostomatal setae, pcx. palpcoxal setae. The definition of cingulum follows Athias-Henriot 1977, as a sclerotized strip which fuses the peritrematal shield with the anterolateral corner of the opisthogastric shield at the level of coxa IV.
The material is deposited in the Canadian National Collection (CNC) of Insects and Arachnids, Ottawa and the Museum d'Histoire Naturelle (MHNG), Geneva, Switzerland (one male, two females of O. lindquisti and one female paratype of O. makarovae).
Taxonomy
The new genus has a peculiar combination of features which make it unique in the subfamily Pergamasinae (Juvara-Balş 2002; Hrúzová and Fend'a (2018). The main characteristics to define a new genus in this subfamily are:
Both sexes: The position of peritrematal shield in connection with the dorsal and ventral one; the presence or absence of the cingulum (Athias-Henriot 1977); the chaetotaxy of dorsal and ventral idiosoma, and the number of gland-pores, especially the presence or absence of gland-pore gv1; the shape of the gnathotectum and the form of the palpfemoral and palpgenual setae al. Female: the number of teeth on movable digit of chelicera and the characteristics of the epi- and endogynium.
The new genus shares with Paragamasus Hull 1918 sensu lato (Athias-Henriot 1971, Karg 1993, Hrúzová and Fend'a 2018) the following characteristics: the number of dorsal setae and the leg chaetotaxy (Evans 1963), the peritrematal shield, which is free posteriorly and attached anteriorly to dorsal shield, the trifid gnathotectum and the spatulate setae al1, al2 inserted on palpgenu. The form of dorsal setae s1, s2 (very tiny) and those of male's genital lamina, rounded with a concavity at its anterior margin, resemble those described (Micherdzinski 1969, Juvara-Balş 1977) in Aclerogamasus Athias-Henriot, 1971, a former subgenus of Paragamasus sensu lato.
The new genus Occigamasus differs from the other genera of Pergamasinae in having four teeth on movable digit of female chelicera and the peritrematal shield free posteriorly. This characteristic is found only in Heteroparasitus (Medioparasitus) athiasae (Juvara-Balș 2002) but this genus is different in many other features. Witalinski (1972) noted a variation of three or four teeth on the movable digit in Heteroparasitus quadratus Witalinski, 1972. The fact that some species have four teeth seems to be an allopatric characteristic. Other characteristics which characterize Occigamasus are the pilose seta inserted on dorsal face of fixed digit as well as the palpfemoral seta al pectinate with four tines (O. lindquisti n. sp.) or more in the other related species. These features are otherwise specific to some genera of the subfamily Parasitinae.
On podonotum all the setae of the dorsal hexagon (z5, j5, and j6) are similar in length and width. In Occigamasus n. gen. seta pd2 is very long on tarsus IV. This characteristic is homoplastic, being described also in some species of Paragamasus, Thenargamasus, Leptogamasus and even in the subfamily Parasitinae (Athias-Henriot 1969a, Micherdzinski 1969, Hrúzová and Fend'a 2018).
Descriptions
Occigamasus new genus
ZOOBANK: 06A18EF2-963A-4E10-AE3C-AF4E8C3E808E ![]()
Figs. 1–9
Type species — Occigamasus lindquisti n. sp.
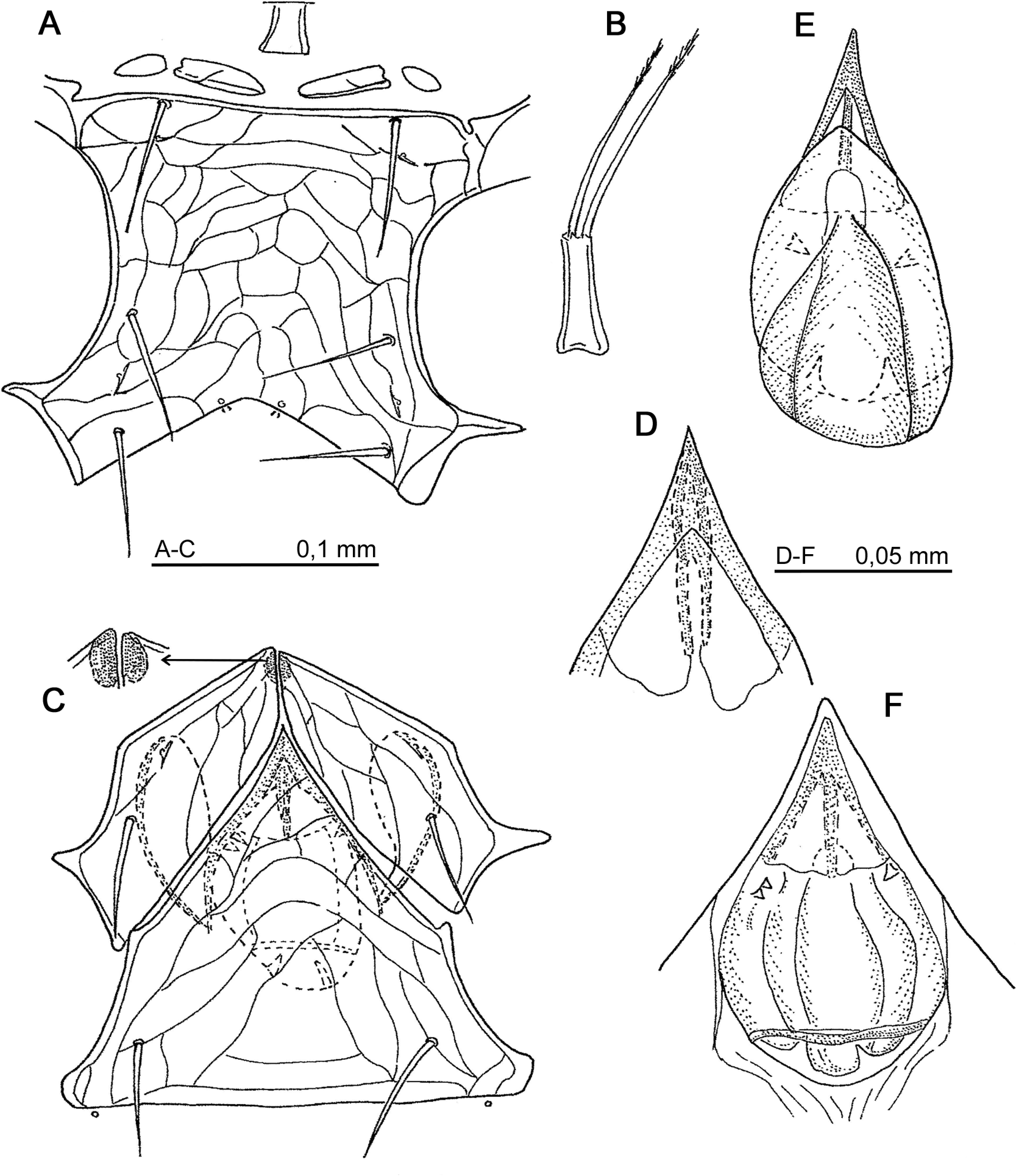
Diagnosis — Dorsal shield of adult entire in female, with a median transverse suture between the podonotum and opisthonotum in male. Podonotum with 22 pairs of acicular, simple setae, slightly curved; setae s1 and s2 tiny (6–14). Opisthonotal region with 20–21 acicular, simple setae, their tips reaching the following row of setae. Dorsal adenotaxy with five gland pores: gdj2, gdz5, gds4, gdz6, gdZ3. Ventral adenotaxy with three pairs of gland pores: gv1 and gv3 with one solenostome, gv2 with two solenostomes; gv1 located closed to medial line of posterior margin of sternal shield or in vicinity of seta st3. Epigynium subapical structure spindle-like. Endogynium formed as a big sack which has various structures inside. Ventrianal shield with eight pairs of opisthogastric setae, ZV1 very short (6–7), JV5 inserted on soft cuticle. Peritrematal shield free from ventrianal shield posteriorly and attached to podonotal region of holodorsal shield anteriorly.
Gnathotectum simply trifid. Hypognathal groove with 10–11 denticulate rows. Palpgenu with setae al1, al2 spatulate; palpfemoral seta al pectinate with four or more tines. Movable digit of female chelicera with four teeth, movable digit of male with two teeth; dorsal seta on fixed digit pilose. Trochanter IV, dorsally, with a small protuberance bearing a tiny spine at its apex (Fig. 5E).
Etymology — The name of the genus combines the Latin suffix ''occi''-, from, ''occidens – occido'' meaning `west', with - ''gamasus'', referring to gamasine mites. It means the gamasus from the west.
Occigamasus lindquisti n. sp.
ZOOBANK: C72F41AF-161A-48F9-8413-EB6A549EE64F ![]()
Figs. 1–6


Type material — Canada. British Columbia (B. C.), Vancouver Island: Holotype: 1 female, Cowichan Lake, Experimental Station, mixed Douglas fir, alder, fern litter, 25 July 1975. Paratypes: 2 females, 1 male, same data as holotype; Cowichan Lake Experimental Station, Mesachie Lake: 1 male, 2 females, Douglas fir litter, 24 July 1975; 1 female, from moss, herbs on log; 3 females, moss on rock; 1 male, 2 females, from mixed cedar deciduous shrub litter, 25 July 1975; 1 male, from alder, crab apple litter, 8 July 1975.
Other material examined. Canada. Vancouver Island (B. C.): 2 males, 2 females, Miracle Beach Provincial Park, from Douglas fir shrub litter, 10 July 1975. U.S.A., Oregon Curry County: 3 males, 3 females, Loeb State Park, 8 miles NE, from tan oak, willow, fern litter, 12 July 1985.
Diagnosis — Length of idiosoma: 936–940 female, 768–816 male. Both sexes: gnathotectum with three simple prongs, the central one slightly shorter than the lateral ones. Female: epigynial spindle-like structure 72 long; endogynium a double sack with two curved prongs on its posterior margin and one or two denticles on lateral walls; sternal shield with gland pores gv1 located medially on posterior margin; on tarsus IV seta pd2, 240 long. Male: armature of leg II with simple rounded protuberance on genu, triangular spur on tibia and a basal protuberance on femoral apophysis; on tarsus IV seta pd2, 190–216 long.
Etymology — The species is dedicated to the eminent acarologist Evert E. Lindquist who collected the mites and gave us the opportunity to study the material.
Female (n = 16)
Figs. 1 A–C F, 2–5
Dorsal idiosoma (Figs. 1A, B) — Colour brownish-yellow, holodorsal shield ornamentation finely reticulate on podonotum, scutellate on opisthonotum. Length of idiosoma 936–940. Peritrematal shield united anteriorly with dorsal shield and free posteriorly; length of peritreme ca. 360. All dorsal setae acicular and simple.
Podonotum with 22 pairs of setae, their length: j1 = 36; j2 = 84–96; j3, j4, j5 = 108–114; j6 = 78; z1 = 24; z2 = 36; z3 = 42; z4 = 84; z5 = 60; z6 = 96–108; s1 = 12, s2 = 6–7; s3 = 96–100; s4 = 60; s5 = 120; s6 = 40; r2 = 36; r3 = 168–180; r4 = 30; r5 = 36.
Opisthonotum with 20–21 pairs of simple setae, slightly curved, their tips reaching alveoli of following row of setae; their length from 84 to 108. On lateral soft integument five pairs of UR setae with lengths 6, 8–9, 18, 36 and 54, respectively. A tiny, fine seta, x, (6–12), probably a conversion of a poroid, located on soft cuticle posteriorly to peritrematal shield. Dorsal adenotaxy with five gland-pores: gdj2, gdz5, gds4, gdz6, gdZ3. Poroidiotaxy with five pores on podonotum and seven on opisthonotum.
Ventral idiosoma — Tritosternum located between presternal plates, its length = 54, basal width = 30; laciniae pilose, 120–132 long (Fig. 2B). Presternal plates formed by two pairs of sclerites, inner one long ellipsoidal and outer one shorter and round; sternal shield reticulated, its mid-length 132–144; gland pores gv1 located centrally on posterior margin; distance between gv1–gv1 = 36 (Fig. 2A). Distance between pairs of sternal setae: st1–st1 = 120; st2–st2 = 120–144; st3–st3 = 132–144. Length of sternal setae: st1 = 60; st2 = 66–72; st3 = 78. Paragynia weakly reticulated with bean-like, sclerotized formations on their anteromedial margin, st4 = 66; metagynial sclerite ellipsoidal; epigynium weakly reticulated, its subapical structure spindle-like with a posteriorly rounded membrane (Figs. 2C, D). Epigynium length (h) = 192–216, basal width (b) = 228–240; h/b = 0.8-0.9; length of st5 = 54, genital pores iv5 near posterior margin on soft cuticle. Endogynium (Figs. 2E, F) a big sack extending dorsally with a pocket-like structure; on its lateral margin with one or two teeth; posterior margin with two curved prongs (12 long) and distance between them from 18 to 36; this variation depends on different localities from which females were sampled. Ventrianal shield (Fig. 1F) with eight pairs of opisthogastric setae, their lengths: ZV1 = 7, ZV2 = 96, ZV3 = 48, ZV4 = 72, JV1 = 48, JV2, JV4 = 66; JV3 = 77. Setae JV5 = 72 on soft cuticle. Gland pore gv2 with two solenostomes, gland pore gv3 simple near anal opening. Length of simple paranal setae 24 and of anal seta 12.
Gnathosoma — Gnathotectum simple trifid, central prong slightly shorter than lateral ones (Fig. 3C). Hypognathal groove with 10 rows of fine denticles; palpcoxal setae pilose, hypostomatic setae simple, their lengths: hyp.1, pcx = 72; hyp.2, hyp.3 = 65. Corniculi triangular; palptrochanter with simple setae v1 (42) and pilose setae v2 (48) (Fig. 3A). Palpfemur with seta al quadripectinate and palpgenu with setae al1, al2 entire, spatulate (Fig. 3B). Chelicera (Figs. 3D, E): movable digit with four teeth; fixed digit with two teeth distal to pilus dentilis and a pilose seta on dorsal face.
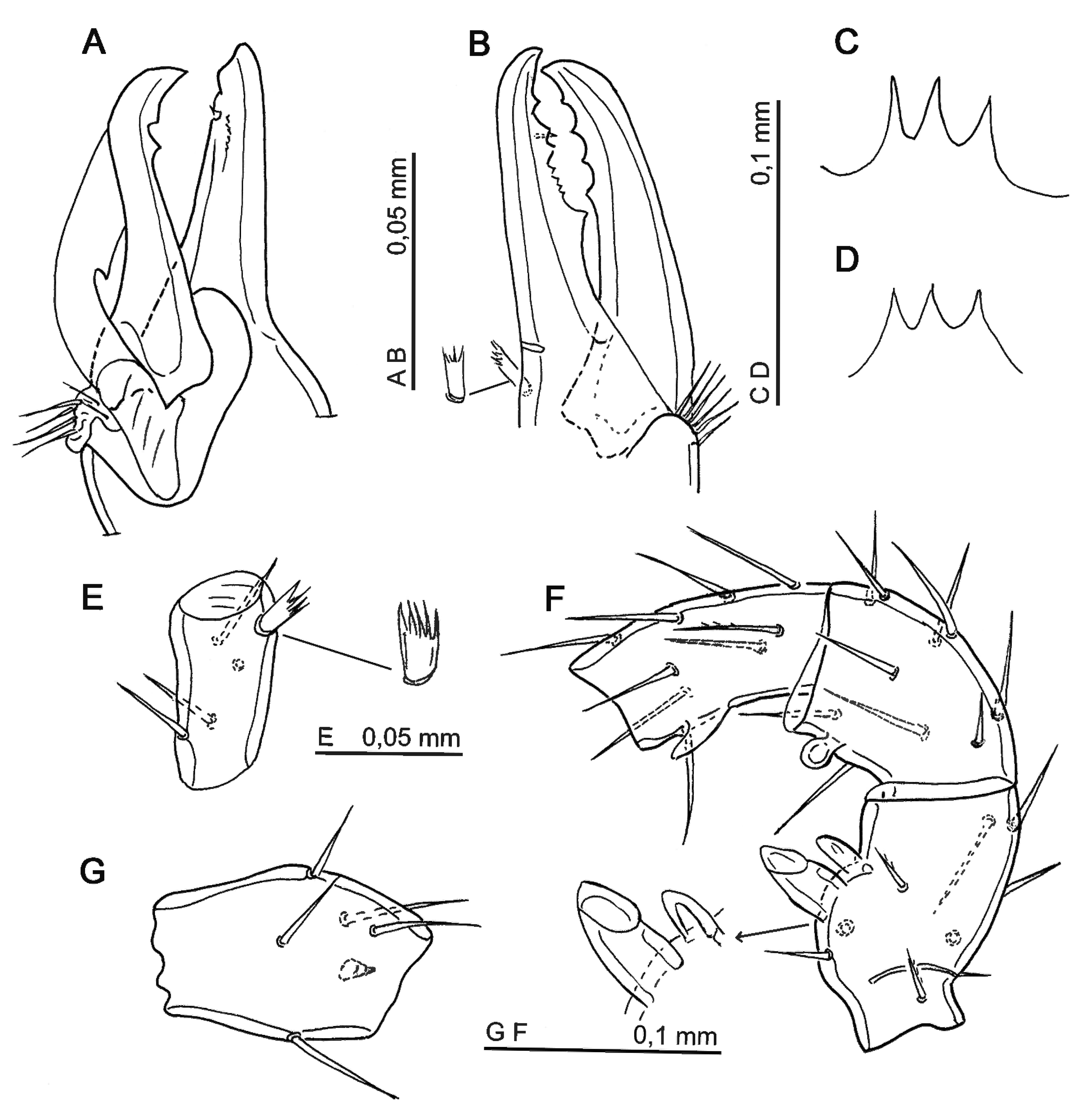
Legs (Figs. 4, 5) — Chaetotaxy, specific to Parasitidae as established by Evans (1963) for genus Pergamasus. Leg lengths: I = 1056 (tarsus = 250–252); II = 770–787; III = 630; IV = 1128 (tarsus = 360–372).


Chaetome : coxae I-IV: 2-2-2-1; trochanter I-IV: 6-5-5-5; femur: I = 13 (2-5/4-2); II = 11 (2-2/2 3/1-1); III = 6 (1-4/1-0); IV = 6 (1-4/1-0); genu: I = 13 (2-3/2 3/1-2); II = 11 (2- 3/1 2/1-2); III = 9 (2 2/1 2/1 1); IV = 10 (2-2/1 3/1-1); tibiae: I = 14 (2-3/2 3/2-2); II = 10 (2-2/1 2/1-2); III = 8 (2-1/1 2/1-1); IV = 10 (2-1/1 3/1-2); tarsi II–IV: 18.
On tarsus (leg II–IV) setae av1, pv1 short (18) and stout; al1, pl1 setae fine (48 long), other setae long and simple or slightly pilose.
Setae on leg I mostly simple, thin, their lengths around 66–72, except av (genu) short and pilose, pd3 (tibia) simple, 36 long. Tarsal dorso-distal sensory field, located in the distal third of this segment, consists of a capsule with an arched phanere 48 (sensilla s Lindquist and Moraza 2016) and three solenidia (24–30), followed by 10 long sensilla inserted subapically; a single solenidion located ventrally. Leg II (Figs. 4A, C) most of setae simple, their lengths from 72 to 84; on tarsus md simple (96), mv stout and short; av1, pv1 short (12), al1, pl1 (48); setae al2, al3, av2, pl3, pd3 slightly pilose. Tarsus III (Fig. 4D) with long (102) setae al3, pl3, mv stout and short (36), other setae simple or slightly pilose (pl2, al2, ad3). Tarsus IV (Fig. 5A) elongated, with long simple setae; some slightly pilose (al3, pl3, ad3, pd3). Seta pd2 very long (240) almost as long as telotarsus. Lengths of some other setae: pl3 = 84; al3, ad3 = 120; mv = 96; al2 = 115; pl2, av2 = 130; pv2 = 72; al1 = 48; ad1, pd1 = 18. A strip of desclerotization present subapically at level of setal insertion av2 on telotarsus (Fig. 5A). Tibia, genu, femur (Figs. 5B–D) with long fine setae. Trochanter IV with a dorsal protuberance bearing a tiny spine apically (Fig. 5E).
Male (n = 9)
Figs. 1 D–E, 3 F, 6

Idiosoma 768–816 long (Figs. 1D–E). Holodorsal shield provided with a transverse suture, which delineates slightly reticulate podonotum from scutellate opisthonotum. Peritrematal shield 330–324 long. Chaetotaxy, adeno- and poroidotaxy as in female. Lengths of some simple podonotal setae: j1 = 54, j2 = 60, j3 = 96–108, j5 = 100, j6 = 78; z1 = 24–30, z2 = 24, z4 = 84, z5 = 60, z6 = 84; s1-s2 = 6, s4 = 90, s5 = 84–90, s6 = 72; r2 = 36–48, r3 = 144–168, r4 = 18, r5 = 78. Opisthonotum with simple, acicular, curved setae, their tips reaching the following row of setae; their lengths 60-72.
Ventral idiosoma — Slightly sclerotized, cuticular ornamentation reticulate on sternogenital shield and scutellate on opisthogaster. Tritosternum as in female, base length 60, width 36; laciniae 78–84 pilose. Presternal platelets ovoid, each with a tiny anterior fragment. Genital lamina rounded, anterior margin slightly concave; subgenital microsclerite rectangular with rounded anterolateral corners (Fig. 6B). Setae in sterno-genital part simples, their length: st1, st3 = 72; st2 = 66; st4 = 66; st5 = 54. Gland pore gv1 simple, distance between gv1–gv1 = 50; gland pore gv2 double with two solenostomes.
Opisthogaster with eight pairs of ventral setae: ZV1 = 6, others from 36 (ZV2) to 84 (JV5). Length of simple paranal setae 24 and of postanal seta 18. Gland pore gv3 near paranal setae.
Gnathosoma — Gnathotectum simple trifid, all prongs of same length (Fig. 3F). Hypognathum and corniculi as in female. Hypognathum with 10 rows of denticles; length of setae: hyp.1 = 54, hyp.2 = 48, hyp.3 = 36–42; slightly pilose pcx. = 60. Palptrochanter with setae v1 simple and v2 pilose. Palpfemur and palpgenu as in female. Chelicera (Fig. 6A): fixed digit slightly hooked, with subapical denticle distal to pilus dentilis and a row of tiny denticles behind it; dorsal seta pilose. Movable digit with two teeth; arm of spermatotreme with inner protuberance in proximal third; arthrodial membrane fan shaped as a range of setule.
Legs — Lengths of legs: I = 1024 (tarsus = 247–252); II = 756; III = 688; IV = 1044–1128 (tarsus = 336–360). Chaetotaxy of legs as in female. Armature of leg II illustrated in Figs. 6C, D: pyramidal femoral apophysis with small protuberance at it base and short thumb-like axillary process; rounded protuberance on genual distal margin; triangular apophysis located medially on tibia. Leg IV: seta pd2, on tarsus, from 190 (site Miracle Beach) to 216 (site Cowichan); trochanter, dorso-laterally, with a protuberance with a spine apically.
See remarks under O. makarovae n. sp.
Occigamasus makarovae n. sp.
ZOOBANK: 638F2793-AC16-4702-8BBE-A758AC1B6020 ![]()
Figs. 7 C–D, 8

Type material — U.S.A., Oregon, Deschutes County: 1 female holotype; 1 female, 2 males, 2 deutonymphs, paratypes. Proxy Falls, St. NWY, 28 miles SW Sisters, ca. 1000m alt., from wet moss and deep damp mixed maple-hemlock litter, 19 July 1985.
Diagnosis — Female endogynium a double sack with a median finger-like protrusion and a membrane with two anterolateral denticles; spindle-like epigynial structure long (66–72). Male femoral apophysis thumb-like.
Etymology — The species is dedicated to Olga Makarova for her major contributions to systematics of Gamasina families, including Parasitidae.
Description — Only the characteristics specific to O. makarovae are described. Chaetotaxy, adenotaxy and poroidotaxy as in the new genus.
Female (n = 2)
Figs. 7 C–D, 8 B–C, E
Dorsal idiosoma — Length of idiosoma 864; length of peritrematal shield 336. Lengths of some simple acicular, setae: on podonotum :j1 = 48, j2 = 84, j3–j5 = 108; z1 = 18–24, z2 = 42; s1 = 18, s2 = 12; r2 = 24, r3 = 168; others from 96 to 108; on opisthonotum length of slightly curved setae from 96 to 108.
Ventral idiosoma — Characteristics of sternal shield, paragynia, metagynial sclerite, epigynium as well as their dimensions as in O. lindquisti. Gland pore gv1 located medially, distance gv1–gv1 = 24. Endogynium (Fig. 7C) a big sack with a median finger-like protrusion on its posterior margin and a membrane with two anterolateral denticles; epigynium with a long (66–72) subapical spindle-like structure surrounded by a triangular membrane with its posterior margin sclerotized (Figs. 7C, D). Ventrianal shield with 8 pairs of setae, their length: ZV1 = 8; ZV2–ZV4 = 60; JV1, JV2, JV4 = 72; JV3 = 66. Setae JV5 = 78 on soft cuticle.
Gnathosoma — Gnathotectum (Fig. 8C) with three smooth prongs their tips at same level. Hypostomatic and palpcoxal setae simple, their length hyp.1, hyp.2, pcx = 72; hyp.3 = 48. Seta al on palpfemur pentapectinate (Fig. 8E). Chelicera (Fig. 8B): movable digit with four teeth; fixed digit with two small teeth between apex and pilus dentilis and behind it a denticulate ridge ended with a tooth; dorsal seta pilose.
Legs — Chaetotaxy typical of Occigamasus n. gen. Setae on legs slender and simple; pd2 on tarsus IV = 228–230. Trochanter IV with dorsal protuberance with a tiny spine apically (Fig. 8G). Measurements: tarsus I = 240; tarsus IV = 372–384.
Male (n = 2)
Figs. 8 A, D, F–G
Lengths of some dorsal setae: j1 = 60, j2, j3 = 100; z1 = 18; s1, s2 = 14; r2 = 60, r3 = 168, r4 = 24; on opisthonotum simple, slightly curved setae, their lengths from 72 to 96.
Ventral idiosoma — Genital lamina rounded with subgenital microsclerite rectangular with anterolateral corners rounded. Other characteristics as in O. lindquisti.
Gnathosoma — Gnathotectum smoothly trifid, all prongs of same length (Fig. 8D). Chelicera (Fig. 8A): fixed digit straight with slightly truncate apex, inner margin with a ridge of small denticles behind pilus dentilis; movable digit as in O. lindquisti.
Legs — Chaetotaxy as in female. Armature of leg II illustrated in Fig. 8F: femoral apophysis thumb-like, axillary process ellipsoidal and straight. Spurs on genu and tibia as in O. lindquisti. Measurements: tarsus I = 240; tarsus IV = 336. Length of seta pd2 (tarsus IV) = 204.
Deutonymph (n = 2)
Fig. 9
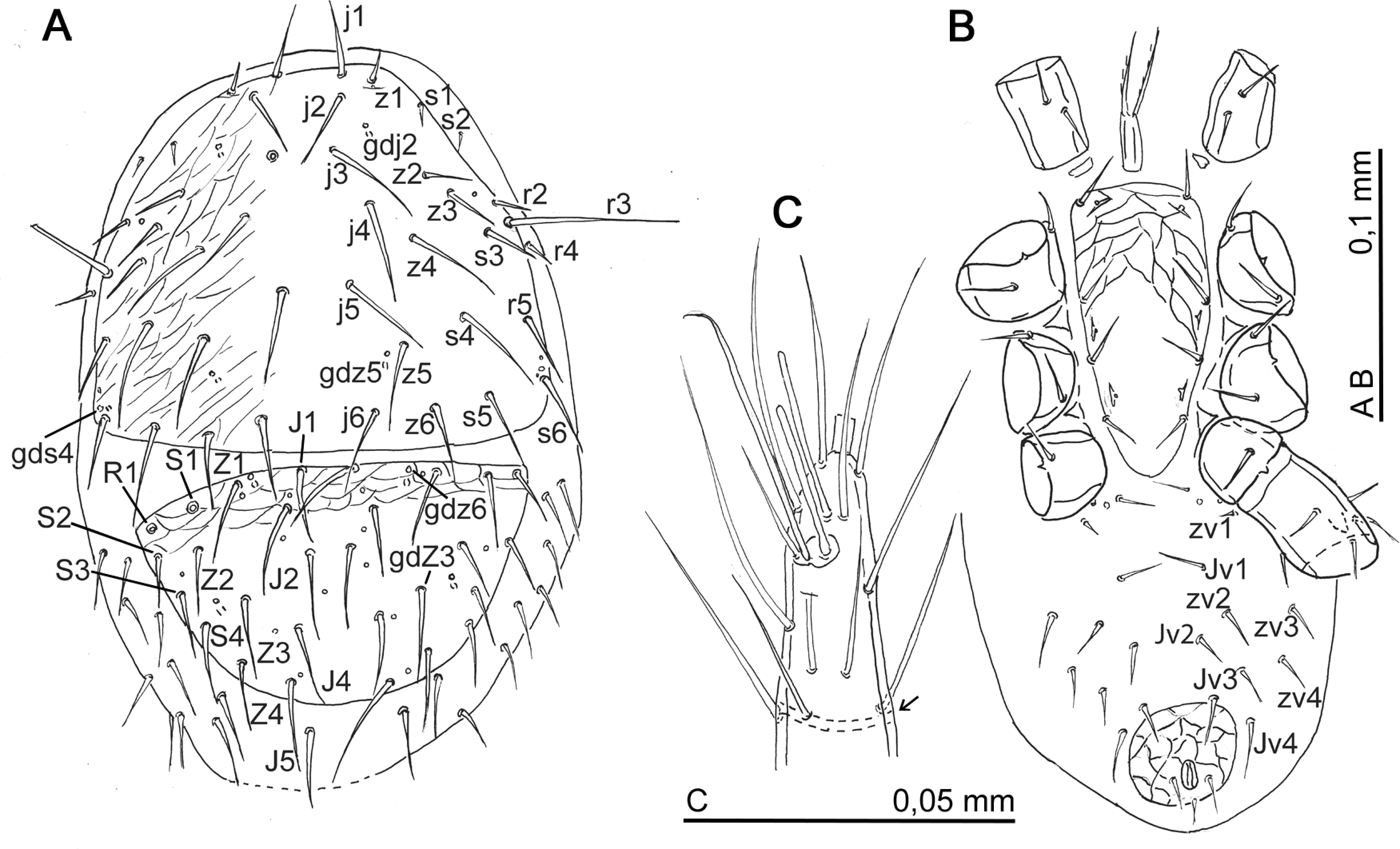
Idiosoma. Weakly sclerotized, yellowish colored. Podonotal shield reticulated only laterally while opisthonotal, sternal, and anal shields well reticulated. All setae simple. Peritrema as in adult.
Dorsal idiosoma (Fig. 9A) — Podonotal shield: mid-length 360, width 380 (measured at its widest point). Podosoma with 20 pairs on podonotal shield and two pairs (s2 and r4) inserted, off the shield, on soft cuticle. Short setae: z1 = 14–15; s1 = 6–10, s2 = 12; r2 = 30, r4 = 18. Other setae from 60 to 96. Setae r3 very long 145–156. Adenotaxie: gland pores gdj2, gdj5, gds4 weakly developed. Gland pore gdz6 shifted on anterior margin of opisthonotal shield. Opisthonotal shield with 14 pairs of setae, their length from 72 to 96, seven pairs of pores and gland pores gdZ3. Only setae R1 inserted on shield, other setae of R series and submarginal setae (UR) located on soft cuticle.
Ventral idiosoma (Fig. 9B) — Tritosternum flanked by a pair of small platelets. Sternal shield with four pairs of setae (st1–st4) and with three pairs of of lyrifissures iv1–iv3. Setae st5 simple, lyrifissures iv5 near coxae IV. Gland pore gv2 not developed. Opisthogastric region with eight pairs of setae. Setae ZV1 very small (2–3). Only pores ivo4 and ivp visible.
Gnathosoma — All the characteristics as in female.
Legs — Chaetotaxy as in adult. Leg I has, apically, a strip of desclerotization located ventrally behind the sensory field (Fig. 9C). Measurements: tarsus I = 240; tarsus IV = 300; setae pd2 on tarsus IV = 204–216.
Remarks — I could not observe all the characteristics of the deutonymph. The two specimens were squashed so that the posterior part of the idiosoma was in bad condition and the chelicerae were hidden into the gnathosoma. However, I remarked two peculiar characters. On the podosoma the setae s2 and r4 were inserted outside the shield like in Parasitinae (Hyatt, 1980). The seta R1 is located on the opisthonotal shield as in Pergamasinae (Bhattacharyya, 1963). The leg I has apically a ventral fissure which was not mentioned in Pergamasinae. Probably this strip of desclerotization helps the movement of tarsus I which is a sensory segment.
The most important feature distinguishing the female of O. makarovae from O. lindquisti is the shape of the endogynium (Figs. 2E, 7C). The posterior margin of the endogynium has in O. lindquisti two curved prongs about 12 long while in O. makarovae only a finger-like protrusion. The fixed digit of the chelicera of these two species is different. Both species have two denticles between pilus dentilis and apex. The inner margin of O. lindquisti, which follows the denticles, is smooth whereas in O. makarovae, behind the pilus dentilis, there is a denticulate ridge ended by a tooth. The male of O. makarovae is similar to those of O. lindquisti but it is recognizable by the characteristics of femoral apophysis on leg II and by those of the fixed chelicera. In O. makarovae the femoral apophysis is thumb-like whereas O. lindquisti has a protuberance on its base. The fixed digit of chelicera in O. makarovae has a truncate apex and those of O. lindquisti a slightly hooked one.
Undescribed species of Occigamasus new genus and their sites
In the Canadian National Collection of Acari (CNC) I identified material of other undescribed species belonging to the new genus. Unfortunately, there was not enough material to describe the new taxa properly. The specimens are mounted dorsally or ventrally so that not all characteristics could be observed. The males are absent in some of the samples. To facilitate future systematic studies, I will mention the sites from where they were sampled, the characteristics of the female endogynium which separate these species (Fig. 7) and some morphological features. Unless otherwise indicated, the samples were collected by E. E. Lindquist.
Occigamasus sp. 1
Material examined — U.S.A. Oregon, Benton County: 1 male, 1 female Mc Glynn Corvallis, ravine, moss on bank, 23 Jan. 1977, coll. L. Russel.
Morphological characteristics — Female (Figs. 7A, B): endogynium a sack with a denticulate plate; epigynium dorsally, with long, sclerotized subapical structure; gv1 posterad st3. Male: leg II with a short apophysis on genu and an elongate spur on tibia; trochanter IV without protuberance. Measurements. Female: tarsus I = 204; tarsus IV = 300, seta pd2 = 120.
Occigamasus sp. 2
Material examined — Canada. British Columbia, Vancouver Island: 1 female, NE intersection Loc Hwy 4 – 45° lat., cedar litter nearby bog, 29 July 1975; idem, 2 females, Heather Mt. trail, 10 miles NW Caycuse, 1000 m alt., moss in creek, 4 Aug. 1975.
Morphological characteristics — Female (Fig. 7E): endogynium with two lateral processes and a denticulate medial plate; epigynium with the sclerotized spindle-like structure (60 long); gv1 located medially on posterior margin of sternal shield. Trochanter IV, dorso-laterally, with a spine apically on a protuberance. Measurements: tarsus I = 264; tarsus IV = 384–420; seta pd2 = 252.
Occigamasus sp. 3
Material examined — Canada, Alberta: 1 female, 1 male, Waterton Lakes National Park, edge of Rowe Lake, from litter, alpine exposure, 2500 m alt., 2–5 Sept. 1980; 1 female, rocky slopes, damp moss; 1 female, 1 male, from grass and substrate, subalpine, by Lake Rowe, 1950 m alt.
Morphological characteristics — Female (Fig. 7F): endogynium a sack with medial spine, lateral protuberances and four anterolateral denticles on the upper margin; epigynium with spindle-like structure; gv1 located medially on posterior margin of sternal shield. Trochanter IV, dorso-laterally, with triangular protuberance. Male: armature on leg II with a small rounded protuberance on genu and trapezoidal spur on tibia. Measurements. Female: tarsus I = 240, tarsus IV = 360; seta pd2 = 216. Male: tarsus I = 228, tarsus IV = 288; seta pd2 = 192.
Occigamasus sp. 4
Material examined — Canada. British Columbia, Vancouver Island: 2 females, Heather Mt. trail, 10 miles NW Caycuse, from hemlock-huckleberry litter, 1000 m, 4 Aug. 1975; 1 female, 1 deutonymph, Comox Glacier, mature hemlock, fir, spruce forest, 176 m alt., moist litter (AF 384), 27 Aug. 1983, coll. A. Fjellberg.
Morphological characteristics —Female (Fig. 7G): endogynium a big sack ventrally with a bifid structure having denticles between the branches, on the anterior margin with a denticulate club surrounded by big denticles; epigynium, dorsally, with a sclerotized ellipsoidal structure with a central spine; gv1 located lateral to seta st3; distance gv1–gv1 = 144. Trochanter IV, dorso-laterally, a small protuberance with a tiny spine apically. Measurements. Female: tarsus I = 252–288, tarsus IV = 336–384; seta pd2 = 156 (Heather Mt Trail) to 240 (Comox Glacier).
Occigamasus sp. 5
Material examined — Canada. British Columbia, Vancouver Island: 4 females, SE end Long Beach, from salal-kinni-kinnick (Arctostaphylos uva-ursi L.)litter on sand dunes, 29 July 1975.
Morphological characteristics — Big mite (Idiosoma length = 1104–1152), well sclerotized, colour brown-reddish. Female (Figs. 7I, J): on sternal shield gv1 in vicinity of st3; endogynium a sack with a club structure anteriorly and many large spines on lateral walls; on posterior margin with a sclerotized, ellipsoidal structure. Epigynium with a bifid, slightly sclerotized subapical structure (Fig. 7K). Trochanter IV with small triangular protuberance on its dorso-lateral face. Measurements. Female: tarsus I = 288, tarsus IV = 396–420; seta pd2 = 288.
Discussion
The geographical distribution of Occigamasus is, until now, limited to the west coast of North America i.e., Vancouver Island, Oregon and California which are included in the Oregonian biotic province and to the Waterton Lakes National Park, Alberta which belong to the Montanian biotic province (Dice 1943 p.128 in Scudder 1979). A preliminary examination of additional material containing the undescribed species 1–5 suggests that in the future we will be able to divide Occigamasus into several species groups. Therefore, for a better understanding of this genus and its species diversity, to find and examine additional specimens from more localities in northwestern North America.
The form of setae on the legs, especially those of tarsus IV and tarsus I, the latter dorso-distal sensory field, has been thoroughly examined only in some genera of Parasitidae (Athias-Henriot 1969a, 1980; Micherdzinsky 1969; Hennessey and Farrier 1989; Al-Atawi et al. 2002). On tarsus I the number and form of solenidia differ among the genera of Pergamasinae (Micherdzinsky 1969, Juvara-Balş unpublished data). For example, Occigamasus has fewer solenidia when compared with Aclerogamasus (Juvara-Balş 1977). The leg chaetotaxy in the whole family needs a revision in order to evaluate the importance of this characteristic in taxonomy.
Another problem brought up during this study, is placement of the species Cycetogamasus californicus (Banks). This species was described as Gamasus californicus by Banks (1904), then transferred by Essig (1958) to the genus Parasitus and subsequently included by Hennessey and Farrier (1988) in the genus Cycetogamasus Athias 1980, subfamily Parasitinae. The genus Cycetogamasus established by Athias-Henriot (1980) belongs to the'' neogamasidienne series'' (série néogamasidienne Athias-Henriot 1977, 1980) which is a different lineage from Pergamasus, Parasitus or even Neogamasus. The main characteristics of females of this genus are the separation of the podonotum and opisthonotum, presence of the cingulum, setae ZV1 very small, and the movable digit with three teeth. Athias-Henriot (1980) added four other species to the genus Cycetogamasus and specified ''je range dans ce nouveaux taxon une série, probablement hétéroclite d'espèces ''néogamasidienne'' (1980 pg. 289). The female of C. californicus differs from those species by the following characteristics: absence of the cingulum, four teeth on the movable digit of the chelicera and dorsal shield not separated, only with a partial transverse split behind setae z6 (Figs. 17, 18, lectotype in Hennesey and Farrier 1988). The common features of female C. californicus with Occigamasus are: scutum peritrematal free posteriorly; on podonotum, 20–22 pairs of acicular setae, s1 and s2 tiny; 22 pairs of acicular setae on opisthonotum; opisthogaster with ZV1 tiny and gv2 double, eight pairs of opisthogastric setae; movable digit of female with four teeth; palpgenual al setae entire and palpfemoral al seta pectinate.
I conclude that C. californicus must be included in the new genus Occigamasus which belongs to the subfamily Pergamasinae Juvara-Balş.
Occigamasus californicus (Banks, 1904) n. comb.
Gamasus californicus Banks, 1904: p. 368
Parasitus californicus Essig, 1958: p. 35
Cycetogamasus californicus (Banks, 1904) Hennessey and Farrier, 1988 (lectotype designated: p. 8).
Another species which should be reviewed is Neogamasus sakhalinus Volonikhina, 1993. This species cannot be included in the genus Neogamasus, as defined by Athias-Henriot 1977, for the following reasons: lack of a cingulum in the female and two teeth on male's movable digit, versus one in Neogamasus. The description and figures are not very detailed and need to be augmented with the characteristics of legs chaetotaxy as well as those of the dorsal and ventral adenotaxy. N. sakhalinus belongs to the ''neogamasidien series''sensu Athias-Henriot 1980, with genus placement yet to be determined.
The mites included in this heterogeneous group need more research in order to redefine the genus or subgenus categories.
I have had the opportunity to study other material, undescribed species, from the CNC from western and eastern Canada and also from Virginia State, U.S.A. The Parasitidae from North and South Carolina have been identified by Hennessey and Farrier (1989). There are great differences between the specimens from the east and those from the west. The species collected from the eastern Canada and northeastern U.S.A. are more similar with those of Europe. Hennesey and Farrier found in Carolina soils Psilogamasus hurlbutti Athias-Henriot (1969b) (subfamily Parasitinae) which was collected from Tanzania and Phytiogamasus primitivus (Oudemans,1904) (subfamily Pergamasinae) which is a Gondwanian species dispersed in the southern hemisphere (Juvara-Balş and Athias-Henriot 1972). At present the paucity of information on the systematics of Parasitidae mites does not permit a discussion of the relationships between Occigamasus of northwestern North America and other genera found on the east coast of Asia (Athias-Henriot 1977, 1980; Tseng 1995). Probably their closest relatives may be found on the east coast of Asia.
Acknowledgements
I would like to thank Evert. E. Lindquist for access to specimens of the Canadian National Collection and for the useful remarks and corrections for the manuscript. For reviewing the manuscript and helpful comments I kindly thank Kamila Hrúzová. I'm indebted to an anonymous reviewer for valuable suggestions. I thank Christina Lehmann-Graber, Gilles Roth (MHNG) for scanning the illustrations and Jean Mariaux and the Department of Invertebrates (MHNG) for providing working space and laboratory facilities.
References
Al-Atawi F., Klompen H., Moser J. C. 2002. Redescription of Schizosthetus lyriformis (McGraw and Farrier.1969) (Parasitiformes: Parasitidae), with revision of the genus. Internat. J. Acarol. 28 (4): 341-360. doi:10.1080/01647950208684310 ![]()
Athias-Henriot C. 1967a. Observations sur les Pergamasus. I. sous-genre Paragamasus Hull, 1918 (Acariens anactinotriches: Parasitidae). Mém. Mus. Hist. nat., Sér. A (Zool.) 49 (1): 1-198.
Athias-Henriot C. 1967b. Observations sur les Pergamasus. II. Sous-genre Pergamasus Berlese, 1904 (Acariens anactinotriches: Parasitidae). Acarologia 9 (4): 669-761.
Athias-Henriot C. 1967c. Observations sur les Pergamasus III. Sous-genre Amblygamasus Berlese, 1903 (Acariens Anactinotriches : Parasitidae). Acarologia 9 :762-800.
Athias-Henriot C. 1969a. Les organes cuticulaires senoriels et glandulaires. Poroidotaxie et adénotaxie. B. Soc. Zool. Fr., 94 : 485-492.
Athias-Henriot C. 1969b. Psilogamasus hurbutti n. g. n. sp. gamaside nouveau de Tanzanie (Acariens Anactinotriches, Parasitidae). Ann. Soc. Ent.Fr. (N.S), 5 (2): 439-449.
Athias-Henriot C. 1971. Paragamasus (Tanygamasus) probsti (Oudemans) (systématique, géographie) avec quelques mises au point synonymiques (Arachnides, Gamasides tocospermique, Parasitidae). Zool. Meded. (Leyden) 45 (16): 167-179.
Athias-Henriot C. 1977. Untersuchungen über die Gattung Neogamasus Tikhomirov mit zwölf neuen Arten aus Korea (cari, Parasitiformes, Gamasida). Anls. Hist.-Nat. Mus. Nat. Hung., 69: 311-341.
Athias-Henriot C. 1980. Parasitidae nord-coréens (Parasitiformes) : apparentés au genre Neogamasus Tikh., avec la création de trois nouveaux genres. Anls. Hist.-Nat. Mus. Nat. Hung. 72: 285-294.
Banks N. 1904. Some Arachnida from California. Proceedings of the California Academy of Sciences. Zoology 3 (13): 331-374.
Banks N. 1910. New American mites. Proceedings of the Entomological Society of Washington 12: 2-12, pls. 1-3 doi:10.1093/besa/2.2.12 ![]()
Beaulieu F., Knee W., Nowell V., Schwarzfeld M., Lindo Z., Behan-Pelletier V., Lumley L., Young M.R., Smith I., Proctor H.C., Mironov S.V., Galloway T.D., Walter D.E., Lindquist E.E. 2019. Acari of Canada. ZooKeys 819: 77-168. doi:10.3897/zookeys.819.28307 ![]()
Berlese A. 1906. Monographia del genere Gamasus Latr. Redia 3: 66-304.
Bhattacharyya S.K. 1963. A revision of the British mites of the genus Pergamasus s. lat. (Acari: Mesostigmata). Bull. Br. Mus. Nat. Hist. (Zool.) 11 (2): 131-242. doi:10.5962/bhl.part.4717 ![]()
Essig E.O. 1958. Insects and Mites of Western North America. The Macmillan Company, New York. pp. 1050
Evans G.O. 1963. Observations on the chaetotaxy of the legs in the free-living Gamasina ( Acari. Mesostigmata). Bull. Br. Mus. Nat. History (Zool.) 10: 275-303. doi:10.5962/bhl.part.20528 ![]()
Evans G.O, Till W.M. 1979. Mesostigmata mites of Britain and Ireland (Chelicerata: Acari, Parasitiformes). An introduction to their external morphology and classification. Trans. zool. Soc. Lond., 35: 139-270. doi:10.1111/j.1096-3642.1979.tb00059.x ![]()
Ewing H.E. 1909. New North American Acarina. Trans. Acad. Scien. ST. Lois (Missouri) 18: 53-77, pls. 8-11.
Hennessey M.K., Farrier M.H. 1988. Systematic revision of thirty species of free-living, soil inhabiting Gamasine mites (Acari: Mesostigmata) of North America. N.C. ARS, Tech. Bull. 285: pp. 123.
Hennessey M.K., Farrier M.H. 1989. Mites of the family Parasitidae (Acari: Mesostigmata) inhabiting forest soils of North and South Carolina. N. C. ARS, Tech. Bull. 292: pp.78.
Hrúzová K., Fend'a P. 2018. The family Parasitidae (Acari: Mesostigmata)-history, current problems and challenges. Acarologia 58 (Suppl): 25-42.
Hyatt K.H. 1980. Mites of the subfamily Parasitinae (Mesostigmata: Parasitidae) in the British Isles. Bull. Br. Mus. Nat. Hist. (Zool.), 38 (5): 237-378.
Johnston D. E., Moraza M.L. 1991.The idiosomal adenotaxy and poroidotaxy of Zerconidae (Mesostigmata: Zerconina). In: Dusbabek, F. & Bukva, V. (eds). Modern Acarology. SPB academic, The Hague, 2: 349-356.
Juvara-Balş I. 1972. Mixogamasus, un nouveae genre de Parasitidae (Acariens : Anactinotriches) de Roumanie. Acarologia 14: 3-14.
Juvara-Balş I. 1977. Quelques remarques sur les Aclerogamasus de Roumanie A. bicalliger Athias et A. motasi n. sp. (Acarina : Gamasida Parasitidae). Trav. Inst. Spéol. "Emil Racovitza", 16:37-45.
Juvara-Balş I. 2002. A revision of the genus Heteroparasitus new status, with the description of Heteroparasitus (Medioparasitus) athiasae subgen. n., sp. n. from Spain and with a key to the genera of Pergamasinae (Acari, Gamasida , Parasitidae). Rev. suisse Zool., 109 (19): 23-46. doi:10.5962/bhl.part.79579 ![]()
Juvara-Balș I., Athias-Henriot C. 1972. Définition de Phityogamasus n. g. et redescription de son espèce type, Parasitus primitivus Oudemans (Gamasides, Parasitidae). Acarologia 13: 446-459.
Karg W. 1993. Acari (Acarina), Milben: Parasitiformes (Anactinotricha). Cohors Gamasina Leach. Raubmilben. Tierwelt Dtl., Gustav Fischer Verlag, Jena, 59: 1-475
Lindquist E.E. 1994. Some observations on the chaetotaxy of the caudal body region of gamasinae mites (Acari: Mesostigmata), with a modified notation for some ventro-lateral body setae. Acarologia 35: 323-326.
Lindquist E.E., Evans G.O. 1965. Taxonomic concepts in Ascidae, with a modified setal nomenclature for the idiosoma of Gamasina (Acarina, Mesostigmata). Mem. Entomol. Soc. Canada 47: 1-64. doi:10.4039/entm9747fv ![]()
Lindquist E.E., Aiscough B.D., Clulow F.V., Funk R.C., Marshall V.G., Nesbitt H.H.J., OConnor B.M., Smith I.M., Wilkinson P.R. 1979. Acari. In: Danks H. V. (Ed.) Canada and its Insect Fauna. Mem. Entomol. Soc. Canada 108: 252-290.
Micherdzinski W. 1969. Die Familie Parsitidae Oudemans 1901 (Acarina, Mesostigmata). Kraków: Panstwowe Wydawnictwo Naukowe. pp. 660.
Oudemans A.C. 1901. Notes on Acari. Ser.3. Tijdschr. nederl. dierk. Vereen., Leyden: 50-88.
Oudemans A.C. 1904. Acarologische Aanteekeningen X. Ent. Ber. 1: 140-141. doi:10.5962/bhl.part.1124 ![]()
Richards L.A. 1976. Parasitid Mites associated with bumblebees in Alberta, Canada (Acarina: Parasitidae; Hymenoptera: Apidae). I. Taxonomy. Uni. Kansas Scien. Bull. 50, 13: 731-773.
Scudder G.G.E. 1979. Present patterns in the fauna and flora of Canada. In: Danks H. V. (Eds) Canada and its insect fauna. Mem. Entomol. Soc. Canada 108: 87-179. doi:10.4039/entm111108087-1 ![]()
Tseng Y-H. 1995. A taxonomical study of free-living gamasine mite family Parasitidae Oudemans (Acari, Mesostigmata) from Taiwan. J. Taiwan Mus., 48: 11-81.
Volonikhina I.I. 1993. New species of Gamasid mites of the genus Neogamasus (Parasitiformes, Gamasina) from the South of the Far East. Zool. Zh., 72 (8): 11-21.
Witalinski W. 1972. Mites of the genus Holoparasitus Oudemans, 1936 (Acarina, Parasitidae). Acta Zool. Cracov., 17 (9): 217-238.



2019-03-25
Date accepted:
2019-12-10
Date published:
2019-12-13
Edited by:
Faraji, Farid

This work is licensed under a Creative Commons Attribution 4.0 International License
2019 Juvara-Balş, Ilinca
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)