



Biology of Mononychelus planki (Acari: Tetranychidae) on Calopogonium mucunoides (Plantae: Fabaceae)
Da Cruz, João Boavida1 ; Massaro, Marcela R. da Silva2 and Moraes, Gilberto José de3
1✉ Departamento de Entomologia e Acarologia, Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, 13418-900 Piracicaba, São Paulo, Brazil.
2Departamento de Entomologia e Acarologia, Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, 13418-900 Piracicaba, São Paulo, Brazil.
3Departamento de Entomologia e Acarologia, Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, 13418-900 Piracicaba, São Paulo, Brazil.
2019 - Volume: 59 Issue: 4 pages: 571-576
https://doi.org/10.24349/acarologia/20194350Original research
Keywords
Abstract
Introduction
Mononychellus planki (McGregor) is a tetranychid known in Brazil as the soybean-green-mite, which unlike many tetranychids produces little or no web. The common name refers to the dark green color of the body of the mite. The tip of the aedeagus of the adult male is subtriangular in lateral view (McGregor, 1950).
This species was originally described from Puerto Rico, on Erythrina berteroana Urb. As summarized by Migeon and Dorkeld (2019), it has been reported from North America (United States and Mexico), Central America (Costa Rica and El Salvador), South America (Argentina, Brazil, Colombia, Paraguay and Venezuela) and the Caribbean (Cuba, Guadeloupe, Puerto Rico, Trinidad and Tobago). It has been found on 68 plant species of 12 families. About 76 % of those plants belong to Fabaceae (35 species) and Malvaceae (17 species), whereas other species belong to Compositae, Cucurbitaceae, Euphorbiaceae, Lamiaceae, Moraceae, Phyllanthaceae, Poaceae, Rubiaceae, Solanaceae and Theaceae.
Mononychellus planki has been cited as the main mite in soybean cultivation in Brazil (Guedes et al., 2004; Roggia, 2007, Moraes and Flechtmann, 2008). In a survey carried out by Roggia (2010), M. planki was found attacking soybean in 52 % of the 27 municipalities evaluated in the Brazilian southernmost state of Rio Grande do Sul. It is usually found on both leaf surfaces, turning them yellowish (Vacante, 2015).
Although damage to soybean by this mite increased in Brazil in the first decade of the 21st century (Moraes and Flechtmann, 2008), when an expansion of the area cultivated with glyphosate tolerant transgenic soybeans took place, Guedes et al. (2007) failed to experimentally demonstrate the higher susceptibility of transgenic soybean to M. planki. This mite has also been mentioned attacking other important crops as cotton (Gossypium hirsutum L.), peanut (Arachis hypogaea L.) and okra (Abelmoschus esculentus L.) (Moraes and Flechtmann, 2008; Moura and Guimarães, 2014; Gabriel, 2016).
In a survey we recently conducted in the Brazilian state of São Paulo (unpublished), M. planki was found on leaves of nine spontaneously growing plant species, namely Blainvillea rhomboidea Cass (Asteraceae), Calopogonium mucunoides Desv. (Fabaceae), Chloris barbata Sw. (Poaceae), Commelina benghalensis L. (Commelinaceae), Croton glandulosus L. (Euphorbiaceae), Lolium multiflorum L. (Poaceae), Physalis angulata L. (Solanaceae), Richardia brasiliensis Gomes (Rubiaceae) and Trema michrantha (L.) (Cannabaceae). The methodology adopted did not allow in determining if those plants were hosts for the mite, but based on Migeon and Dorkeld (2019), it has not been previously reported from those plants.
Studies on the biology of M. planki were conducted on soybean genotypes (Roggia, 2010; Siqueira 2011). The objective of the present work was to evaluate the importance of C. mucunoides as a host for M. planki, evaluating its development and reproduction on this plant, which is commonly attacked by this mite in São Paulo state.
Materials and methods
The work was carried out from January to April 2019, with a mite population collected from C. mucunoides on the campus of the University of São Paulo at Piracicaba, São Paulo state.
Each experimental unit consisted of a 1.5 cm diameter disc taken from a mature but not senescent leaf of C. mucunoides. The discs were placed with the top surface up (treatment 1) or down (treatment 2) on a piece of nylon foam mat held in a tray. The performance of the mite was considered on both leaf surfaces, as the mite is found on both under field conditions. The mat was kept permanently soaked with mineral water to maintain disc turgidity and prevent mite escape. In total, 50 units were used for each treatment. To maintain proper physiological quality, discs were replaced every third day. The units were kept in a chamber at 25 ± 1 °C, 70 ± 10 % RH and a 12 h of daily photoperiod.
Initially, a couple (male and female) of M. planki was randomly taken from C. mucunoides leaves and transferred to each unit. Observations were made every 12 h until at least one egg was laid. The couple and eggs were removed, except for one egg, and the units continued to be examined every 12 h to determine the duration of each immature stage. When the female adult stage was reached, a male obtained from the same plants was transferred to each unit. Thereafter, the units were examined once a day to determine the duration of the adult phases (pre-oviposition, oviposition and post-oviposition) and the oviposition rates. The data obtained were compared using Mann–Whitney U test,with R software.
Parameters of fertility life tables - net reproductive rate (R0), intrinsic population growth rate (rm), mean generation time (T) and finite ratio of population growth (λ) - were estimated and compared using TWOSEX-MS Chart program (Chi, 2018) with paired bootstrap tests (p < 0.05). For those calculations, two different sex ratios were considered for mites on the upper leaf surface, given the rather different values determined for the parental population (obtained from eggs laid by field collected females) and for the respective offspring. A single sex ratio value was considered for the lower leaf surface, because of the similar values obtained for both parental and offspring populations.
Results
Development and reproduction
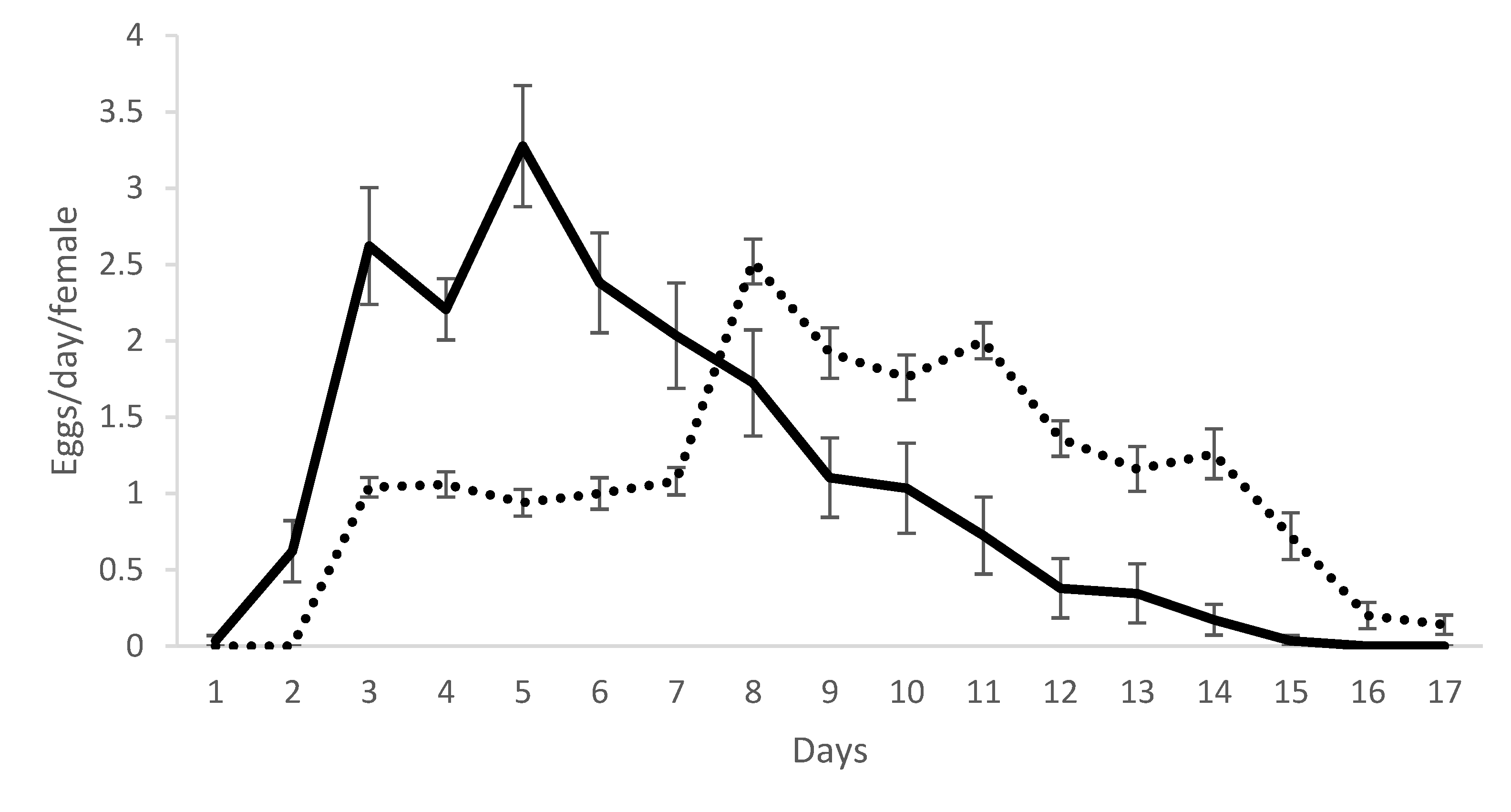
Significant differences were not observed for durations of the larval and protonymphal stages when mites were maintained on either of the two leaf surfaces (Table 1). However, the duration of the egg stage was significantly longer for mites maintained on the lower (5.2 ± 0.2 days) than on the upper (3.3 ± 0.4 days) leaf surfaces, while the opposite was observed for the deutonymphal stage (lower surface: 2.3 ± 0.2 and upper surface: 3.1 ± 1.1 days). As a result, total duration of the immature phase (egg–adult) was statistically similar for mites maintained on the lower (11.9 ± 0.2 days) and upper (11.5 ± 0.9 days) leaf surfaces. Survival of all immature stages was 100 % for mites on the upper leaf surface. For mites on the lower leaf surface, survival varied from 82.9 % for the protonymph to 91.8 % for the egg stage, resulting in 63.3 % survival for the total immature development. Oviposition and post-oviposition periods were longer for mites on the upper leaf surface (12.0 ± 0.3) than on the lower leaf surface (3.0 ± 0.2 days).
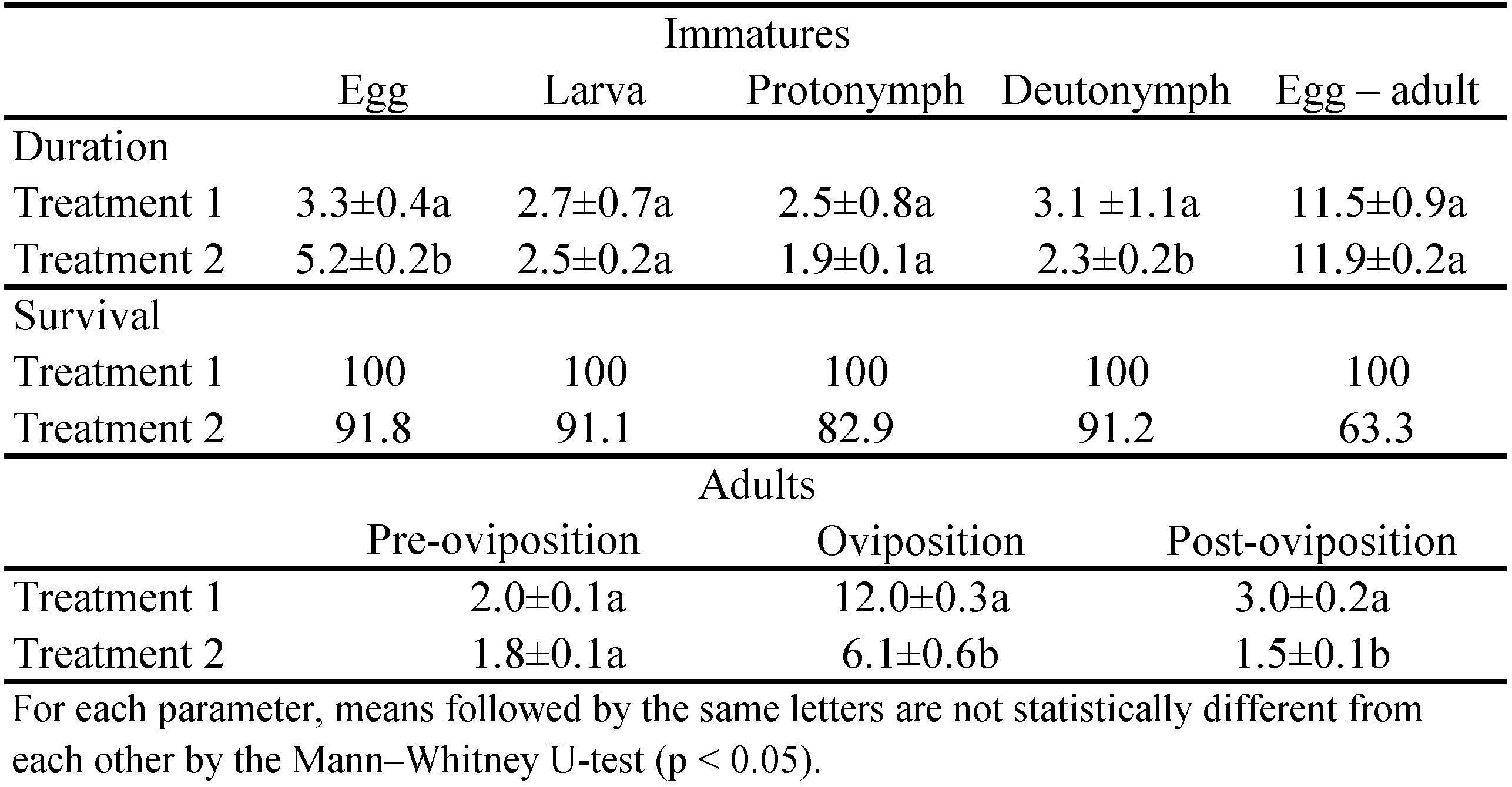

Peak oviposition rates were about the same on both leaf surfaces and occurred earlier for mites on the lower leaf surface (Figure 1). Hence, daily oviposition rate was always higher for mites on the lower surface until the seventh day of oviposition, inverting afterward, as indicated by the non-overlapping 95 % confidence intervals at each day.
Mean fecundities of mites on the upper and lower leaf surfaces were statistically similar (18.1 ± 0.5 and 18.7 ± 2.3 eggs/ female, respectively). These values corresponded to mean daily oviposition of 1.2 ± 0.2 eggs/ female/ day for mites on both leaf surfaces.
Life table
For mites on the upper leaf surface, the parent population (obtained from eggs laid by field collected females) consisted entirely of females, whereas for the offspring, females constituted 53 % of the mites. For this reason, life table was constructed considering the two sex ratios separately, producing, different values for the different parameters (Table 2).

No significant difference between T (mean generation time) values was observed for mites on the upper leaf surface with the different sex ratios, but values of rm, R0 and λ were higher for the population composed only of females. Yet, the values of those parameters in both cases were not high (rm= 0.113 and 0.150 for sex ratios of 53 % and 100 % females), probably because of the low fecundity, as the duration of the immature phase observed is similar to that of other spider mite (less than 12.0 days at about 25 °C) (Crooker, 1985) and survivorship was maximum (100 %).
For mites on the lower leaf surface (Table 2), sex ratio of the parental generation (74 %) was similar to that of the offspring (75 %). In this case, rm, R0 and λ were intermediate to the values reported for mites on the upper leaf surface for the sex-ratio 53 % and 100 % females. T (mean generation time) value was significantly lower (though slightly) for mites on the lower leaf surface than on the upper one (whatever the sex-ratio).
Discussion
The non-significant difference between the duration of the entire immature development of M. planki on both leaf surfaces is consistent with the tendency to find this mite on both sides of the leaves under field conditions. The duration of the immature stages and the different phases of the adult stage are generally comparable to those determined by other authors for this mite species on different soybean varieties (Roggia, 2010; Siqueira, 2011). The only exception concerns the egg stage of mites on the upper surface, which in the present study was considerably shorter than observed by those authors, in which mites were maintained only on the lower leaf surface. We could not find an explanation for this difference, but it could be related to a shorter exposure to UV light, which has been reported as lethal to mites (Suzuki et al., 2009).
On the other hand, fecundity in the present study was very low on both leaf surfaces compared to the values reported by Roggia (2010) and with the values obtained for most of the varieties studied by Siqueira (2011) except for BRS 133, BRS 134, Jackson and PI 200538 varieties, for which fecundity values were comparable to the values obtained in the present study.
The observed differences might be attributed to the intrinsic characteristics of the mite populations used in the study, as the mites studied by Siqueira (2011) were obtained from Londrina, Paraná state. However, this is certainly not the unique factor as mites used by Roggia (2010) were obtained from the same place as the mites of the present study. The physiological quality of the substrate, C. mucunoides and the soybean varieties might also explain the differences. It seems that soybean is a more favorable host, either due to its intrinsic characteristics or due to differences in nutritional quality of the plants as a function of their fertilization in the field. Several studies have demonstrated the best performance of tetranychid mites on plants grown with higher availability of nitrogenous fertilizers (i.e. Alizade et al., 2016).
Regardless these differences, the present study suggests the importance of C. mucunoides in maintaining the population of M. planki in the field under natural conditions, when or where soybean (onto which the biological performance of the mite is better) is not present. Soybean is of Asian origin, having been introduced to Brazil a few decades ago (Mandarino, 2017). Soybean has experienced profound genetic changes due to the intense breeding to which it has been subjected. The increase in yield and other improvements obtained in these works may, in parallel, have made soybean more favorable to the development of certain pests, including M. planki. Concurrently, as M. planki, C. mucunoides is a species native to the American continent (Carvalho-Okano and Leitão Filho, 1985) and commonly found naturally in Brazil. This plant is considered beneficial for several reasons, including its considerable potential for nitrogen fixation and as animal feed (Paulino et al., 2006).
One explanation for the low densities of M. planki on C. mucunoides could the common origin of the two organisms and a possible ''co-evolution''. A complementary or alternative explanation could be the presence of predatory mites of the family Phytoseiidae, which may affect certain phytophagous mites (perhaps also M. planki) in different crops. For example, seven species of predatory mites of the family Phytoseiidae were found on C. mucunoides in observations conducted in nine sites in the state of São Paulo (unpublished data). While collecting specimens of M. planki in the present study, several specimens of the phytoseiid Transeius bellottii (Moraes and Mesa) were collected on the same plant. These predators also may contribute to the maintenance of M. planki at levels that do not cause extreme damage to the host plant and further studies should be performed to confirm this hypothesis.
Acknowledgements
To Ana Clara A.V. B. de Barros and João Pedro G. Silva, for their invaluable help in the conduction of this study. To Prof. Carlos H.W. Flechtmann, for his suggestions in the preparation of this manuscript.
References
Alizade M., Hoseini A., Goldani M., Awal M.M., Hoseini M. 2016. Effects of nitrogen fertilization on population growth of two-spotted spider mite. Syst. & Appl. Acarol., 21: 947-956. doi:10.11158/saa.21.7.8 ![]()
Carvalho-Okano R.M., Leitão Filho H.F. 1985. Revisão do gênero Calopogonium Desv. (Leguminosae - Lotoideae) no Brasil. ver. Brasil. Botânica, 8: 31-46.
Chi H. 2018. TWOSEX-MSCHART. A computer program for age stage, two-sex life table analysis [internet]. [20 June 2018]. Available from http://140.120.197.173/Ecology/ ![]() .
.
Crooker, A. 1985. Embyonic and juvenile development. In: Helle, W. & M.W. Sabelis (eds.). World Crop Pests. Spider Mites. Their Biology, Natural Enemies and Control. Vol. 1A, pp. 149-163.
Gabriel D. 2016. Pragas do amendoim. São Paulo: Secretaria de Agricultura e Abastecimento Agência Paulista de Tecnologia dos Agronegócios. Instituto Biológico. Documento Técnico 26, pp. 25.
Guedes J.V.C., Návia D., Flechtmann C.H.W., Lofego A.C. 2004. Ácaros fitófagos e predadores associadas à soja no Rio Grande do Sul, Brasil. Programas e Resumos do XX Congresso Brasileiro de Entomologia, p. 170.
Guedes J.V.C., Návia D., Lofego A.C., Dequech S.T.B.C. 2007. Ácaros associados à cultura da soja no Rio Grande do Sul, Brasil. Neotropical Entomol., 3: 288-293. doi:10.1590/S1519-566X2007000200017 ![]()
Mandarino J.M.G. 2017. Origem e a história da soja no Brasil. Blog da Embrapa Soja. A Soja no Contexto do Agronegócio Brasileiro. p. 1.
McGregor E.A. 1950. Mites of the family Tetranychidae. American Midland Naturalist, 44: 257-420. doi:10.2307/2421963 ![]()
Migeon A., Dorkeld F. 2019. Spider Mites Web: a comprehensive database for the Tetranychidae [internet] [assessed on 10 May 2019] Montpellier: INRA/CBGP. Available from: http://www1.montpellier.inra.fr/CBGP/spmweb/ ![]() .
.
Moraes G.J., Flechtmann C.H.W. 2008. Manual de acarologia. Acarologia básica e ácaros de plantas cultivadas no Brasil. Ribeirão Preto: Holos Editora. pp. 288.
Moura A.P., Guimarães J.A. 2014. Manejo de pragas na cultura do quiabeiro. Circular Técnica 138. Brasília: MAPA. EMBRAPA. pp. 12.
Paulino V.T., Gerdes L., Valarini M.J., Júnior E.F. 2006. Retrospectiva do uso de leguminosas forrageiras. Encontro Sobre Leguminosas. II Encontro Técnico de Leguminosas Forrageiras. [internet]. [assessed on 10 May 2019] Available from http://www.iz.sp.gov.br/pdfs/1178129826.pdf ![]() .
.
Roggia S. 2007. Ácaros tetraniquídeos (Prostigmata: Tetranychidae) associados à soja no Rio Grande do Sul: ocorrência, identificação de espécies e efeito de cultivares e de plantas daninhas [MSc dissertation]. Universidade Federal de Santa Maria, Santa Maria, Brazil. pp. 113.
Roggia S. 2010. Caracterização de fatores determinantes dos aumentos populacionais de ácaros tetraniquídeos em soja [PhD thesis]. Escola Superior de Agricultura Luiz de Queiroz, Universidade de São Paulo, Brazil. pp. 155.
Siqueira F. 2011. Biologia e flutuação populacional de Mononychelus planki (McGregor) (Acari: Tetranychidae) em cultivares de soja Glycine max (L.) Merr. e impacto de imidacloprido em aspectos biológicos do adulto [PhD thesis]. Universidade Federal do Paraná, Brazil. pp. 79.
Suzuki T., Watanabe M., Takeda M. 2009. UV tolerance in the two-spotted spider mite, Tetranychus urticae. J. Insect Physiol., 55(7): 649-654. doi:10.1016/j.jinsphys.2009.04.005 ![]()
Vacante V. 2015. The handbook of mites of economic plants. Identification, bio-ecology and control. Wallingford: CABI. pp. 876. doi:10.1079/9781845939946.0001 ![]()



2019-05-12
Date accepted:
2019-11-12
Date published:
2019-11-19
Edited by:
Tixier, Marie-Stéphane

This work is licensed under a Creative Commons Attribution 4.0 International License
2019 Da Cruz, João Boavida; Massaro, Marcela R. da Silva and Moraes, Gilberto José de
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)