




Contribution to the knowledge of the oribatid mite genus Eremella (Acari, Oribatida, Eremellidae)
Ermilov, Sergey G.1 and Frolov, Andrey V.2
1✉ Tyumen State University, Tyumen, Russia.
2Zoological Institute of Russian Academy of Sciences, Saint Petersburg, Russia.
2019 - Volume: 59 Issue: 2 pages: 214-226
https://doi.org/10.24349/acarologia/20194326ZooBank LSID: 9EBAAF87-A45E-4C72-B138-AB3CBB0610D4
Original research
Keywords
Abstract
Introduction
The oribatid mite genus Eremella (Acari, Oribatida, Eremellidae) was proposed by Berlese (1913) with Eremella vestita Berlese, 1913 as type species. According to the Subías's checklist (2019), it comprises seven species, which are distributed in the Tropical, Subtropical and southern Palaearctic regions.
In the course of the study of scarab beetles inhabiting termite nests in Brazil, numerous specimens of oribatid mites were found in a nest of Cornitermes silvestrii Emerson in Snyder, 1949 (Isoptera, Termitidae). Examination of the mites showed that they belong to a new species of Eremella. Earlier, only one species of this genus was recorded in the Brazilian fauna, Eremella cf. induta Berlese, 1913, but this record was based exclusively on unconfirmed identification (Oliveira et al. 2005). This paper aims to describe and illustrate the new species, update generic diagnosis, give an identification key to known species of the genus, and discuss the relationship of eremellid genera Eremella with Archeremella Balogh and Mahunka, 1974, Licnocepheus Woolley, 1969, Triteremella Kunst, 1971 and Afreremella Mahunka, 1973.
This work is part of our continuing study of oribatid mites inhabiting termite nests of the world (e.g., Ermilov et al. 2017a, b; Bayartogtokh et al. 2018), and oribatid mite fauna of Brazil (e.g., Ermilov et al. 2014, 2015; Ermilov & Tolstikov 2015).
Methods
Specimens were mounted in lactic acid on temporary cavity slides for measurement and illustration of the new species. Body length was measured in lateral view, from the tip of the rostrum to the posterior edge of the notogaster. Notogastral width refers to the maximum width of notogaster in dorsal view. Lengths of body setae were measured in lateral aspect. All body measurements are presented in micrometers. Formulas for leg setation are given in parentheses according to the sequence trochanter–femur–genu–tibia–tarsus (famulus included). Formulas for leg solenidia are given in square brackets according to the sequence genu–tibia–tarsus.
Drawings were made with a camera lucida using a Leica transmission light microscope ``Leica DM 2500''. Microscope images were obtained with an AxioCam ICc3 camera using a Carl Zeiss transmission light microscope ``Axio Lab.A1''. SEM photos were made with the aid of a JEOL–JSM-6510LV SEM microscope.
General morphological terminology used in this paper mostly follows that of F. Grandjean: see Travé and Vachon (1975) for references, Norton (1977) for leg setal nomenclature, and Norton and Behan-Pelletier (2009) for overview.
The following abbreviations are used: cos = costula; ro, le, in, bs, ex = rostral, lamellar, interlamellar, bothridial and exobothridial setae, respectively; bo = bothridium; c, la, lm, lp, h, p = notogastral setae; im, ip, ih, ips = notogastral lyrifissures; gla = opisthonotal gland opening; a, m, h = subcapitular setae; or = adoral seta; v, l, d, cm, acm, ul, sul, vt, lt = palp setae; sac = axillary saccule; ω = palp and leg solenidion; cha, chb = cheliceral setae; Tg = Trägårdh's organ; Pd I, Pd II = pedotecta I and II, respectively; 1a, 1b, 1c, 2a, 3a, 3b, 4a, 4b = epimeral setae; dis = discidium; g, ag, an, ad = genital, aggenital, anal and adanal setae, respectively; 1, 2, 3 = genital papillae; ψ, τ = setae of ovipositor; iad = adanal lyrifissure; p.o. = preanal organ; Tr, Fe, Ge, Ti, Ta = leg trochanter, femur, genu, tibia and tarsus, respectively; p.a. = porose area; σ, φ = leg solenidia; ɛ = leg famulus; v, ev, bv, l, d, ft, tc, it, p, u, a, s, pv, pl = leg setae.
Systematics
Generic diagnosis of Eremella Berlese, 1913
Adult — With character states of Eremellidae (Balogh 1961; Norton and Behan-Pelletier 2009). Body size: Small (length less than 350). Integument: Heavily areolate-reticulate or tuberculate, often additionally microgranulate. Prodorsum: Rostrum rounded. Costulae of medium size or long, simple, parallel. Transcostula and tutorial ridges developed or not developed. Rostral, lamellar and interlamellar setae short, ro setiform, le and in setiform or phylliform; ro and le inserted nearly to rostrum, in in interbothridial region. Bothridial setae of medium size or long, clavate or flabelliform. Bothridia cup-shaped, with thick margins. Notogaster: Anterior and posterior parts of notogaster rounded. Pteromorphs, lenticulus and octotaxic system absent. With 10 (exception: nine) pairs of notogastral setae setiform or phylliform; dorsal setae c, la, lm, lp, h1–h3 located submarginally, p1–p3 in posterior position. Gnathosoma: Subcapitulum diarthric. Palps with setation 0–2–1–3–9(+ω). Solenidion of palptarsi bacilliform, attached to acm. Axillary saccule present. Lateral podosomal and epimeral regions: Pedotecta I and II represented by small laminae, rounded distally. Genal teeth, custodia and circumpedal carinae absent. Discidium developed or absent. Epimeral setal formula: 3-1-2(or 3)-2(or 3). Anogenital region: Six (rarely five) pairs of genital, one pair of aggenital, two pairs of anal and three pairs of adanal setae. Legs: All legs monodactylous or heterotridactylous. Tibia I with dorsodistal apophysis bearing solenidion φ1. Porose areas present on all femora.
Juvenile instars — Not known.
Eremella vazdemelloi sp. nov.
ZOOBANK: 034FC270-6CA6-414A-B2DA-E694C483CFAA ![]()
(Figures 1–6)
Diagnosis — Body size: 232–265 × 132–157. Body surface densely tuberculate and microgranulate. Costulae long. Tutorial ridges absent. Rostral, lamellar, interlamellar and exobothridial setae short, setiform, barbed; le thickest. Bothridial setae of medium size, with slightly flattened, clavate, barbed head, always directed posterolaterally or posteriorly. Notogastral setae short, setiform, with curved tip, minutely barbed. Epimeral and anogenital setae short, setiform, minutely barbed. Six pairs of genital setae.
Description
Measurements — Body length: 249 (holotype: female), 232–265 (47 paratypes: 32 females and 15 males); notogaster width: 149 (holotype), 132–157 (47 paratypes). No clear difference between females and males in body size.

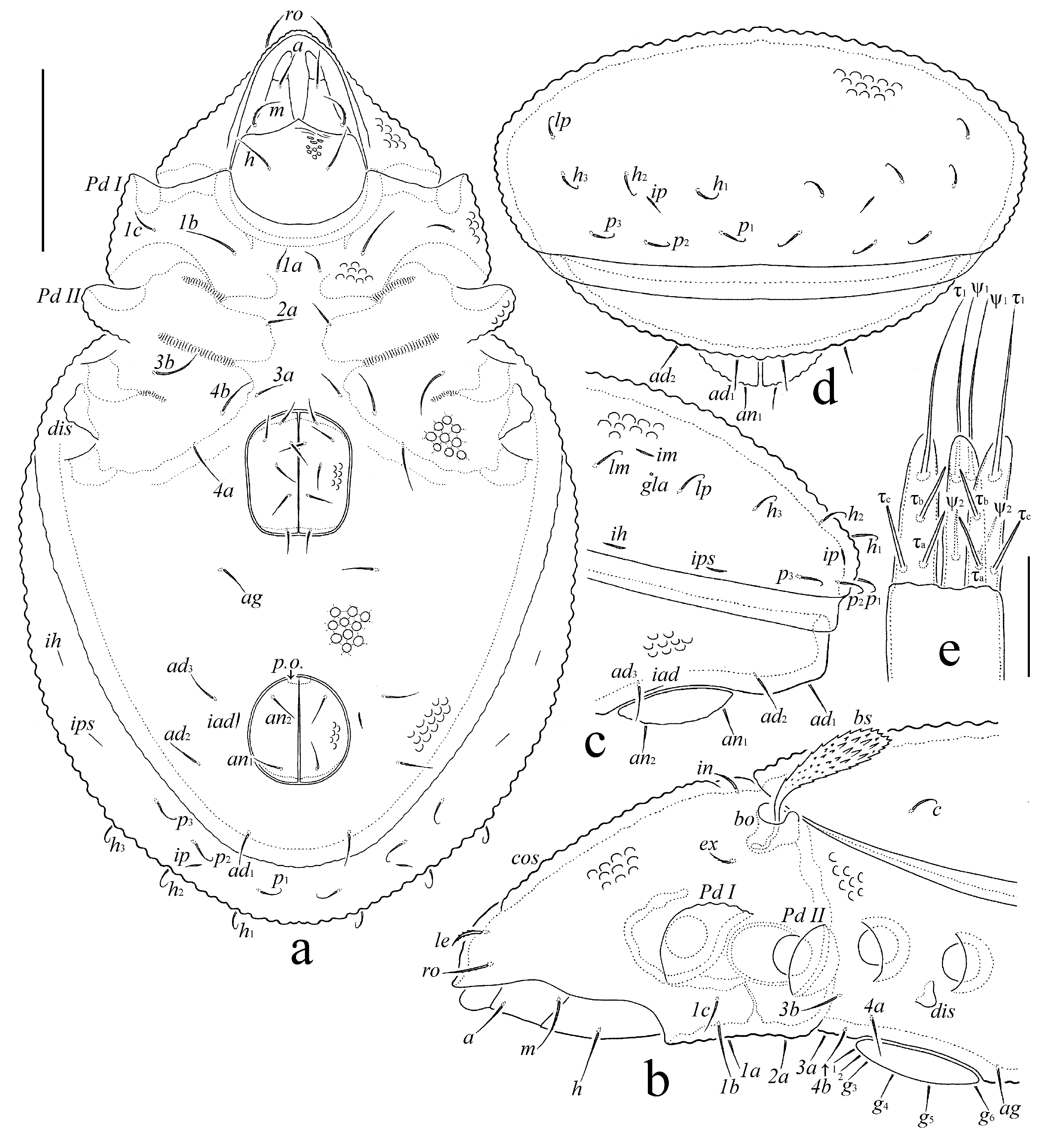



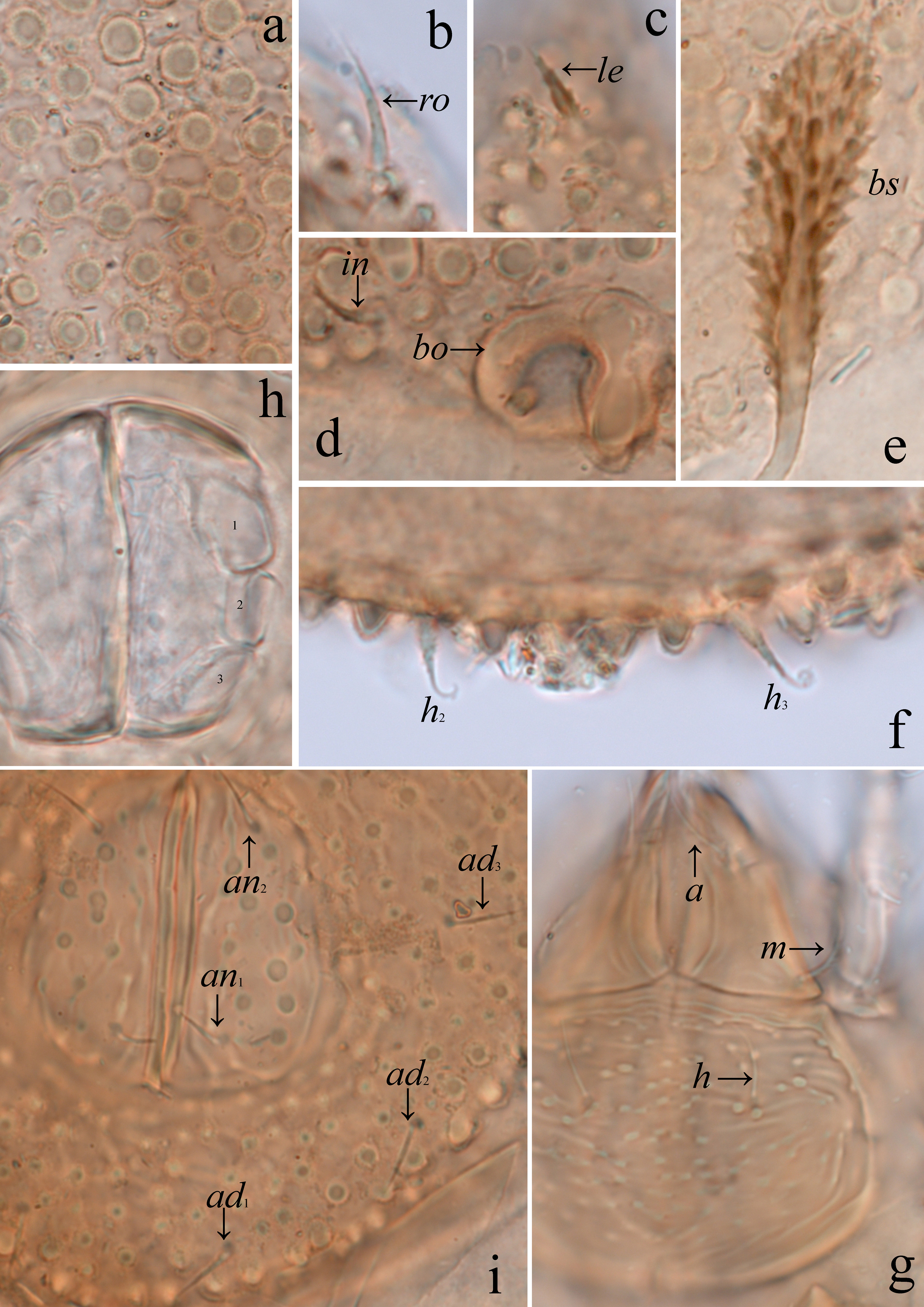
Integument — (Figs 1a, b, 2a–d, 3a–d, 4a–d, 5a–c, 6a, g, i). Body light brown. Body surface (including genital and anal plates) and all leg femora and trochanters III, IV with dense tubercles (their diameter up to 6), which are covered by numerous microgranules (their diameter less than 1); tubercles often connected by slightly developed ridges. Subcapitular mentum with tubercles and striae. Leg genua, tibiae and tarsi with dorsal and paraxial ridges.
Prodorsum — (Figs 1a, 2b, 4a–d, 6b–e). Rostrum broadly rounded. Costulae long, usually distinctly developed (Fig. 4a), rarely poorly visible (Fig. 4b), located dorsally between rostral setae and bothridia. Tutorial ridges completely absent. Rostral (14–16), lamellar (10–12), interlamellar (8–10) and exobothridial (8–10) setae setiform, barbed; le thickest, ex thinnest. Bothridial setae (41–45) with short stalk and well developed, slightly flattened, clavate, barbed head, specifically directed posterolaterally or posteriorly. Bothridia with lateral thickening. Dorsophragmata and pleurophragmata not observed.
Notogaster — (Figs 1a, 2a–d, 4a–c, 6f). Ten pairs of notogastral setae similar in length (10–12), setiform, with curved tip, hardly barbed. Opisthonotal gland openings, lyrifissures (except ia – not found), circumgastric scissure and circumgastric sigillar band poorly visible.
Gnathosoma — (Figs 1b–d, 2a, b, 6g). Subcapitulum longer than wide (53–57 × 36–41). Three pairs of subcapitular setae setiform; a (10–12) and h (14–16) hardly barbed, m (14–16) distinctly barbed. Two pairs of adoral setae (4–6) setiform, smooth. Palps (41–45) with setation 0-2-1-3-9(+ω). Postpalpal setae (2) thorn-like, smooth. Axillary saccules poorly visible, elongated. Chelicerae (57–61) with two setiform, barbed setae, cha (18–20) longer than chb (10–12). Trägårdh's organ of chelicerae tapered.
Epimeral and lateral podosomal regions — (Figs 2a, b). Epimeral setal formula 3-1-2-2. Setae setiform, hardly barbed; 1b (18–20) longer than 3b, 4a, 4b (18) and 1a, 1c, 2a, 3a (10–12). Discidia broadly triangular.
Anogenital region — (Figs 2a–e, 6h, i). Six pairs of genital, one pair of aggenital, two pairs of anal and three pairs of adanal setae similar in length (10–12), setiform, hardly barbed. Adanal lyrifissures located close and parallel to anal plates. Preanal organ small, slightly elongate, conical. Ovipositor elongated (length 65–71, width of cylindrical part 20), blades (24–26) shorter than length of distal section (beyond middle fold; 41–45). Each of the three blades with four smooth setae, ψ1 ≈ τ1 (30–32) setiform, ψ2 ≈ τa ≈ τb ≈ τc (10–12) thorn-like. Coronal setae not observed.
Legs — (Figs 3a–d, 4a–c, 5c, d). Median claw slightly thicker than lateral claws, all strong, smooth. Porose areas on femora I–IV slightly visible, located dorsoparaxially on femora I, II and posteroparaxially on femora III, IV. Porose areas on trochanters not observed. Formulas of leg setation and solenidia: I (1-5-3-4-17) [1-2-2], II (1-5-3-4-15) [1–1–2], III (2-3-1-3-15) [1-1-0], IV (1-2-2-3-12) [0-1-0]; homology of setae and solenidia indicated in Table 1. Famulus of tarsi I thickened, erect, truncate distally, inserted between solenidia. Solenidion φ1 on tibiae I very long, setiform; ω1 and ω2 on tarsi I and φ2 on tibiae I comparatively long, distinctly or slightly thickened, blunt-ended, ω1 specifically curved; other solenidia comparatively short, bacilliform. Dorsoanterior apophysis of tibiae I (bearing both solenidia) well developed.


Material examined — Holotype (female) and 47 paratypes (32 females and 15 males): Brazil, Mato Grosso State, Cuiabá, Flor do Cerrado, 15°29ʹ38″S 56°4ʹ36″W, termite nest (Cornitermes silvestrii) in small patch of Cerradão, a forest-like subtype of the Cerrado, 27.II.2017 (collected by A.V. Frolov).
Type deposition — The holotype and one paratype are deposited in the collection of UNESP—Universidade Estadual Paulista, Department of Zoology and Botany, 15054–000 São José do Rio Preto–SP, Brazil; registration numbers: 1739.1 and 1740.1, respectively. Five paratypes are deposited in the Acarology collection of the Zoological Institute, Russian Academy of Sciences, Saint-Petersburg, Russia. Three paratypes are deposited in the collection of the Senckenberg Institute, Görlitz, Germany. Thirty eight paratypes are deposited in the Tyumen State University Museum of Zoology, Tyumen, Russia. All in ethanol with a drop of glycerol.
Etymology — The new species is named after Dr. Fernando Vaz-de-Mello (Federal University of Mato Grosso, Cuiabá, Brazil), the leading scarab beetle researcher in Brazil.
Remarks — Eremella vazdemelloi sp. nov. is morphologically most similar to Eremella ensifera Balogh and Mahunka, 1968 in having tuberculate body surface and setiform notogastral setae, but differs from the latter by the presence of short notogastral setae (versus medium size), the bothridial setae specifically directed posterolaterally or posteriorly (versus laterally), and the absence of tutorial ridges (versus present).
Discussion
According to the Subías's checklist (2019), besides Eremella the family Eremellidae comprises other genera such as Archeremella, Licnocepheus and Triteremella.
Archeremella (three species) was described as a subgenus of Eremella by Balogh and Mahunka (1974) with type species, Eremella (Archeremella) leowae Balogh and Mahunka, 1974. The genus differs from other representatives of Eremellidae by the presence of 14 pairs of notogastral setae (versus 10 pairs); of these, four pairs inserted in central part of notogaster (versus central part without setae). Based on this difference, Pérez-Íñigo (1981) suggested to raise the status of Archeremella to the generic level, which was supported further by other authors (e.g., Fujikawa 1991; Subías 2004; Mahunka 2008); we also support this opinion.
Licnocepheus (monotypic) was described by Woolley (1969) with type species, Licnocepheus reticulatus Woolley, 1969 in the family Licnodamaeidae, but he did not compare it to the genera of Eremellidae. However, all main morphological traits of L. reticulatus (body surface; morphology of costulae, bothridial setae and ventral side of body; localization of notogastral setae) are similar to those of Eremella, hence there is no distinct difference between these two genera. Therefore, we propose that Licnocepheus as a junior subjective synonym of Eremella, and its known species recombined as follows: Eremella reticulatus (Woolley, 1969) comb. nov.
Triteremella (three species) was described by Kunst (1971) with type species, Eremella kaszabi Csiszár, 1962. This genus is morphologically similar to Eremella, but differs from the latter by the tridactylous legs (versus monodactylous). However, there is another eremellid genus, Archeremella, which included species with one and three claws. Therefore, we propose Triteremella is a junior subjective synonym of Eremella, and following recombination: Eremella simpliseta (Mahunka, 2011) comb. nov. Also, we support the initial placement of species, having tridactylous legs: Eremella kaszabi Csiszár, 1962 and Eremella matildebellae Mahunka and Palacios-Vargas, 1995.
Afreremella (monotypic) was described by Mahunka (1973) with type species, Afreremella luisiae Mahunka, 1973. Subías (2004) synonymized this genus with Triteremella. However, Afreremella differs distinctly from the other eremellid genera by the following important traits: costulae and transcostula forming trapezoid structure (versus costulae and transcostula H-shaped, or transcostula absent); bothridial setae very short, spoon-like (versus medium size or long, clavate to flabelliform). In our opinion, these differences are enough for keeping the status of Afreremella in Eremellidae, therefore we accept the validity of this genus.
Balogh (1959) described Proteremella with type species, Proteremella pulchella Balogh, 1959. This genus differs from Eremella by the presence of setiform dorsal notogastral setae (versus phylliform). Later, Balogh and Mahunka (1974) treated Proteremella as a subgenus of Eremella. Pérez-Íñigo (1981) suggested Proteremella as a junior synonym of Eremella, and he didn't consider the morphology of notogastral setae as generic or subgeneric trait in Eremellidae (for example, Archeremella included species with setiform and phylliform setae).
Key and distribution to known species of Eremella
1. Notogaster with longitudinal median ridge and transverse ridge-like branches
...... 2
— Notogaster without ridges
...... 3
2. Notogaster tuberculate; interlamellar and dorsal notogastral setae setiform; body size: 280 × 150
...... Eremella africana (Balogh, 1966). Distribution: Chad.
— Notogaster areolate; interlamellar and dorsal notogastral setae phylliform; body size: 213–258 × 110–132
...... Eremella matildebellae Mahunka and Palacios-Vargas, 1995. Distribution: Mexico.
3. Notogaster tuberculate
...... 4
— Notogaster areolate
...... 7
4. Dorsal notogastral setae phylliform; body size: 252–271 × 145–147
...... Eremella kaszabi Csiszár, 1962. Distribution: central and southern Europe.
— Dorsal notogastral setae setiform
...... 5
5. Five pairs of genital setae; costulae obviously arch-like, located close to each other in median part; body size: unknown
...... Eremella simpliseta (Mahunka, 2011). Distribution: Madagascar.
- Six pairs of genital setae; costulae slightly arch-like, distinctly removed from each other in median part
...... 6
6. Dorsal notogastral setae of medium size (each seta reaching seta of the following row); bothridial setae directed laterally; tutorial ridges present; body size: 279–311 × 136–168
...... Eremella ensifera (Balogh and Mahunka, 1968). Distribution: Argentina, Iran.
— Dorsal notogastral setae short (each seta clearly not reaching seta of the following row); bothridial setae directed specifically posterolaterally; tutorial ridges absent; body size: 232–265 × 132–157
...... Eremella vazdemelloi sp. nov. Distribution: Brazil.
7. Interlamellar and dorsal notogastral setae phylliform
...... 8
— Interlamellar and dorsal notogastral setae setiform
...... 10
8. Dorsal notogastral setae broadly phylliform, dilated distally; lamellar and notogastral setae of p-serie setiform; bothridial setae with short stalk and large, slightly elongate (almost globular) head; body size: 240–248 × 130–140
...... Eremella induta Berlese, 1913 (see also Balogh & Mahunka 1966; Mahunka & Mahunka-Papp 1995). Distribution: Oriental and Ethiopian regions, Tonga, Japan, “Brazil (?)”.
— Dorsal notogastral setae narrowly phylliform, dilated in median part; lamellar and notogastral setae of p-series slightly phylliform; bothridial setae with long stalk and elongate (clavate or flabelliform) head
...... 9
9. One transcostula developed between costulae; body size: 300 × 180
...... Eremella vestita Berlese, 1913 (see also Mahunka & Mahunka-Papp 1995). Distribution: Oriental region.
— Two transcostulae developed between costulae; body size: 236–264 × 121–150
...... Eremella funnagasatoensis Nakamura, Hashimoto, Nishi, Nakamura and Fujikawa, 2015. Distribution: Japan.
10. Transcostula developed; five pairs of genital setae; body size: 240–275 × 125–146
...... Eremella pulchella (Balogh, 1959). Distribution: central and southern Europe.
— Transcostula not developed; six pairs of genital setae; body size: 186–204 × 102–114
...... Eremella reticulatus (Woolley, 1969). Distribution: U.S.A. (Louisiana).
Acknowledgements
The authors thank Dr. Fernando Vaz-de-Mello (Federal University of Mato Grosso, Cuiabá, Brazil) for logistics support during AVF's field work in Brazil; the Ministério do Meio Ambiente, Brazil, for issuing a collecting permit (SISBIO-50574-1, 11/2015-03/2017); Dr. Julia Baumann (University of Graz, Graz, Austria) and two anonymous reviewers for valuable comments; and Andrey N. Bobylev (Tyumen State University, Tyumen, Russia) for preparing SEM photos. The study was supported by the Russian Foundation for Basic Research (research project №18-04-00096) and in the framework of the State research project АААА-А19-119020690101-6.
References
Balogh J. 1959. Neue Oribatiden aus Ungarn (Acari). Ann. Univ. Sci. Budap., Sect. Biol., 2: 29-35.
Balogh J. 1961. Identification keys of world oribatid (Acari) families and genera. Acta Zool. Acad. Sci. Hung., 7(3-4): 243-344.
Balogh J. 1966. On some oribatid mites from Tshad and East Africa collected by Prof. H. Franz, Vienna. Opusc. Zool. Budapest, 6(1): 69-77.
Balogh J., Mahunka S. 1968. The scientific results of the Hungarian soil zoological expeditions to South America. 5. Acari: Data to the oribatid fauna of the environment of Córdoba, Argentina. Opusc. Zool. Budapest, 8(2): 317-340.
Balogh J., Mahunka S. 1966. The scientific results of the Hungarian soil zoological expedition to the Brazzaville-Congo. 3. The oribatid mites (Acari) of Brazzaville-Congo. 1. Acta Zool. Acad. Sci. Hung., 12(1-2): 25-40.
Balogh J., Mahunka S. 1974. Oribatid species (Acari) from Malaysian soils. Acta Zool. Acad. Sci. Hung., 20(3-4): 243-264.
Bayartogtokh B., Ermilov S.G., Hugo-Coetzee E.A., Khaustov A.A. 2018. Contribution to the knowledge of the oribatid mite genus Licnodamaeus Grandjean, 1931 and synonymy of Licnodamaeolus Covarrubias, 1998 (Acari, Oribatida, Licnodamaeidae). Syst. Appl. Acarol., 23(1): 42-60. doi:10.11158/saa.23.1.4 ![]()
Berlese A. 1913. Acari nuovi. Manipulus 7-8. Redia, 9: 77-111.
Csiszár J. 1962. On an interesting new Eremella species (Acari, Oribatei). Ann. Hist. Nat. Mus. Nat. Hung., 54: 401-403.
Ermilov S.G., Tolstikov A.V. 2015. Additions to the oppioid oribatid mite fauna of Brazil, with description of two new species (Acari, Oribatida, Oppioidea). Biologia, 70(5): 636-644. doi:10.1515/biolog-2015-0072 ![]()
Ermilov S.G., Hugo-Coetzee E.A., Khaustov A.A. 2017a. Coetzeella navalensis gen. nov., sp. nov. (Acari, Oribatida, Oppiidae) from South Africa. Syst. Appl. Acarol., 22(3): 403-409. doi:10.11158/saa.22.3.6 ![]()
Ermilov S.G., Hugo-Coetzee E.A., Khaustov A.A. 2017b. Oribatid mites (Acari, Oribatida) inhabiting nests of the termite Trinervitermes trinervoides (Sjöstedt) in the Franklin Game Reserve (Bloemfontein, South Africa), with description of a new species of the genus Ceratobates (Tegoribatidae). Syst. Appl. Acarol., 22(10): 1715-1732. doi:10.11158/saa.22.10.12 ![]()
Ermilov S.G., Shtanchaeva U.Ya., Bayartogtokh B., Subías L.S. 2015. The oribatid mite genus Lopholiodes (Acari, Oribatida) with description of a new species. Neot. Ent., 44(6): 580-587. doi:10.1007/s13744-015-0310-9 ![]()
Ermilov S.G., Tolstikov A.V., Senna A.R., Pešić V. 2014. A new aquatic species of the oribatid mite genus Mucronothrus (Acari, Oribatida, Trhypochthoniidae) from Brazil. Int. J. Acarol., 40(7): 570-576. doi:10.1080/01647954.2014.962086 ![]()
Fujikawa T. 1991. List of oribatid families and genera of the World. Edaphologia, 46: 1-132.
Kunst M. 1971. Nadkohorta Pancirnini - Oribatei. In: Daniel M., Cerny V. (Eds.), Klic Zvireni CSSR IV. Praha: Academia. pp. 531-580. [in Czechoslovak]
Mahunka S. 1973. Neue und interessante Milben aus dem Genfer Museum XI. Neue und wenig bekannte Oribatiden aus Rhodesien (Acari). Arch. Sci., 26(3): 205-225.
Mahunka S. 2008. A new genus and some other data of oribatids from Thailand (Acari: Oribatida). Acta Zool. Acad. Sci. Hung., 54 (2): 125-150.
Mahunka S. 2011. New and little known oribatid mites from Madagascar (Acari: Oribatida), III. Opusc. Zool. Budapest, 42(1): 43-66.
Mahunka S., Mahunka-Papp L. 1995. The oribatid species described by Berlese (Acari). Budapest: Hungarian Natural History Museum. 325 pp.
Mahunka S., Palacios-Vargas J. 1995. Two new oribatid (Acari: Oribatida) species from the canopy of tropical dry forest in Mexico. Acta Zool. Acad. Sci. Hung., Budapest, 41(4): 295-300.
Nakamura Y.-N., Hashimoto S., Nishi Y., Nakamura Y., Fujikawa T. 2015. Two new species of Eremellidae and Scheloribatidae (Acari, Oribatida) from the Kuma district of southern Japan. Acarologia, 55(2): 171-187. doi:10.1051/acarologia/20152159 ![]()
Norton R.A. 1977. A review of F. Grandjean's system of leg chaetotaxy in the Oribatei (Acari) and its application to the family Damaeidae. In: Dindal D.L. (Ed.). Biology of oribatid mites. Syracuse: SUNY College of Environmental Science and Forestry. pp. 33-61.
Norton R.A., Behan-Pelletier V.M. 2009. Oribatida. Chapter 15. In: Krantz G.W., Walter D.E. (Eds.). A Manual of Acarology. Lubbock: Texas Tech University Press. pp. 430-564.
Oliveira A.R., Norton R.A., Moraes G.J. 2005. Edaphic and plant inhabiting oribatid mites (Acari: Oribatida) from Cerrado and Mata Atlântica ecosystems in the State of São Paulo, southeast Brazil. Zootaxa, 1049: 49-68. doi:10.11646/zootaxa.1049.1.4 ![]()
Pérez-Íñigo С. 1981. Resultados de la expedición Peris-Alvarez a la isla de Annobón (13) Oribatid mites (2nd series). Eos, LVII: 201-212.
Snyder T.E. 1949. Catalog of the termites (Isoptera) of the world. Smithsonian Misc. Coll. Publ., 112(1): 1-490.
Subías L.S. 2004. Listado sistemático, sinonímico y biogeográfico de los ácaros oribátidos (Acariformes: Oribatida) del mundo (excepto fósiles). Graellsia, 60 (número extraordinario): 3-305. doi:10.3989/graellsia.2004.v60.iExtra.218 ![]()
Subías L.S. 2019. Listado sistemático, sinonímico y biogeográfico de los ácaros oribátidos (Acariformes: Oribatida) del mundo (excepto fósiles). Online version accessed in March 2019, 536 pp.; http://bba.bioucm.es/cont/docs/RO\_1.pdf ![]()
Travé J., Vachon M. 1975. François Grandjean. 1882-1975 (Notice biographique et bibliographique). Acarologia, 17(1): 1-19.
Woolley T.A. 1969. A new and phoretic oribatid mite (Acarina: Cryptostigmata: Licnodamaeidae). Proc. Ent. Soc. Washington, 71(4): 476-481.



2019-03-23
Date accepted:
2019-05-27
Date published:
2019-05-29
Edited by:
Baumann, Julia

This work is licensed under a Creative Commons Attribution 4.0 International License
2019 Ermilov, Sergey G. and Frolov, Andrey V.
 Download article Download low definition
Download article Download low definitionDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)