



Tritrophic relationships among tomato cultivars, the rust mite, Aculops lycopersici (Massee) (Eriophyidae), and its predators
Aysan, Elif1 and Kumral, Nabi Alper2
1Department of Plant Protection, Faculty of Agriculture, Bursa Uludag University, 16059, Bursa, Turkey.
2✉ Department of Plant Protection, Faculty of Agriculture, Bursa Uludag University, 16059, Bursa, Turkey.
2018 - Volume: 58 Issue: Suppl pages: 5-17
https://doi.org/10.24349/acarologia/20184283Keywords
Abstract
The tomato rust mite, Aculops lycopersici (Massee) (Acari: Eriophyidae), is a significant pest of cultivated tomato cultivars and feeds on some wild tomato species and other solanaceous species, such as eggplant and nightshade (Mason and Huber 2001; Shipp et al., 2001; Haque and Kawai 2002; Kim et al., 2002; Goldsmith 2004; Ozman-Sullivan and Ocal 2005; Kumral and Cobanoglu 2015a,b). In tomatoes, this mite first causes leaf necrosis, which is similar to a lack of micro-elements in the leaves. High mite populations lead to water stress with smaller, distorted and dusty brown-coloured leaves. When mites feed on the generative organs, inflorescence and young fruit become shrivelled with dropped flowers and russeted fruit, and if uncontrolled, the plants will die (Royalty and Perring 1988; Kumral et al., 2014). In addition, heavy mite infestations lead to crop losses of more than 65% in tomato production (Celar and Valič 2003).
To control these pests in the outdoor tomato fields of Turkey, the standard method is to spray synthetic acaricides based on a calendar date (Anonymous, 2015). Because mites are small, they are difficult to see with a simple magnifier. Chemical treatments are often unsuccessful at controlling these pests as they inaccurately estimate the control time. This method often results in poor mite control and ecological problems, such as destroying non-target organisms and natural enemies of the pests (Yu 2008, Khalighi et al., 2016). Additionally, when the mite population density becomes high, the tomato plant is in its phenological fruit-ripening period. To avoid chemical residue on crops, most growers refrain from using chemicals during this period. Although some biological agents feed on A. lycopersici, they are usually insufficient because the leaves and saps of some plant cultivars are extensively covered with glandular and non-glandular trichomes (Brodeur et al., 1997; Fischer and Mourrut-Salesse 2005; Van Houten et al., 2013a).
The most important factors in the host plant’s favour are type, density and the trichomes’ toxic chemical composition (Van de Boom et al., 2003; Kang et al., 2010). Trichomes are reported to help protect against insects and spider mite attacks (Chatzivasileiadis and Sabelis 1998; Gonçalves et al., 1998; Pocoví et al., 1998; Aragão et al., 2002; Antonious and Snyder 2006; Maluf et al., 2007, 2010; Schie et al., 2007; Alba et al., 2009; McDowell et al., 2011). Glandular and non-glandular trichomes also restrict some predator species from moving on the plant leaf surfaces as well as being toxic and limiting predator development (Kennedy 2003; Buitenhuis et al., 2015). In addition, trichomes can shelter some small pests, protecting them from their natural enemies (Van Houten et al., 2013b).
Such tritrophic interactions may affect A. lycopersici population dynamics in different tomato cultivars. Different cultivars may also reduce the mite’s fitness on the host plant or activate its natural enemies. Consequently, to keep the mite levels below the economic threshold, tomato cultivars that are unsuitable for A. lycopersici, but suitable for its predators, are required. This study determined the population development of A. lycopersici on six tomato hybrid cultivars, namely, Dora, Etna, Grande, H2274, Jana and M1103, under controlled and natural field conditions and evaluated tritrophic interactions among plant trichomes and population densities of A. lycopersici and its predators under field conditions.
Aculops lycopersici were collected from tomato plants on Gorukle Campus (Bursa, Turkey). The species was identified by Eddie A. Ueckermann (Agriculture Research Council, Plant Protection Research Institute, Pretoria, South Africa) based on morphological characteristics viewed under both light and scanning electron microscopes (Kumral et al., 2014). The mite population was mass-reared on potted tomato plants grown in a climate-controlled room. Synchronous mite populations were obtained by rearing at least two generations on each tomato cultivars.
Three stake tomato cultivars, Dora, Etna, and Jana (May-Agro Company, Bursa, Turkey), and three indeterminate hybrid tomato cultivars, Grande, H2274 and M1103, (Agromar Company, Bursa, Turkey) were grown in 2.5-L pots filled with Klasmann TS 1 type peat (Deilmann, Geeste, Germany) in a climate-controlled room on a 16 h light: 8 h dark cycle (27 ± 1°C, 65 ± 5%). Seedlings were irrigated every third day with tap water and fertilized weekly with water-soluble fertilizer containing macro and micronutrients: 3% total nitrogen (N), 7% phosphorus (P2O5), 4.5% potassium (K2O), 0.1% sulphur (SO4.S), 0.25% iron (Fe), 0.01% copper (Cu), 0.1% zinc (Zn), 0.1% manganese (Mn), 0.01% boron (B), and 0.001% molybdenum (Mo) (University of Uludag, Department of Soil Science and Nutrition, Bursa, Turkey). For the population development studies under natural conditions, twenty-five-day-old plants were transferred to a field for sowing. Before sowing, the field was fertilized with only organic manure containing nitrogen. The plants were irrigated at two-day intervals by a drip irrigation system. Tomato seedlings were sprayed with insecticide (40 g/l azdirachtin, Nimiks 4.5, Agrikem Certus, USA) and fungicide (65.82 g/l copper sulphate pentahydrate, Mastercop®M, Bravo Ingeneri Industrial S.A. de C.V., Mexico) during and after the sowing period. For the population development studies under controlled conditions, plants in full bloom with five fully expanded, combined mature leaves of approximately equal area, were used.
To determine population development under controlled conditions, full bloom tomato plants were artificially infested with 25 female mites from the synchronous A. lycopersici population with a soft-bristle paintbrush in 2014. Twenty-one days after infestation, mobile mites were counted on all leaf surfaces and stalks under a stereomicroscope (Leica EZ4, Germany). Each tomato cultivar included three replicates with 3 plants per replicate.
To assess population development under natural conditions, the field experiment was conducted in a 0.1 ha tomato field at the Uludag University Agricultural Faculty in Bursa in 2015. Experimental plots containing 10 plants per plot were constructed based on a Latin square design (Figure 1) with six replicate plots per cultivar. Three leaves were collected at three different heights (top, middle and bottom) from 3 plants per plot. Fifty-four leaves from each cultivar were collected weekly from early June to late October. A. lycopersici mobile stages and their predator mites and insects on the tomato leaves and stalks were counted under a stereomicroscope. Predator mites were preserved in alcohol (70%), then cleaned, stained and mounted in Hoyer’s medium (Hoy 2011). Mounted mite species were identified under an optical microscope per the methods of Baker (1970), Ueckermann & Grout (2007), Faraji et al. (2011). The mite specimens were deposited in Acarology Laboratory of Bursa Uludag University. Based on species identification, weekly mean mite abundance was calculated for each cultivar, and abundance was the number of mites per leaf on each sampling date.


Five leaves were collected from five different levels per tomato when the plants reached the fruit set period under natural conditions. Six 1-mm2 areas on each leaf were examined for trichome density on both the abaxial and adaxial leaf surfaces using a template with different positions for the 1-mm2 areas. Sampled leaves were the same age and position and not selected randomly. Each tomato cultivar included three replicates with 3 plants per replicate. Non-glandular (types II and V) and glandular (type VI) trichomes were identified per the methods of Channarayappa et al. (1992) and McDowell et al. (2011). Trichome density was represented as 1 mm2.
The weekly mean mite population densities were compared among weeks with a repeated measures analysis of variance, followed by Tukey’s multiple comparison test using SAS statistical software (SAS, 2007). One-way analysis of variance ( textitANOVA) was performed on the data from the trichome density and mite population development in controlled conditions, and significant differences among the mean mite numbers and trichome density for the six cultivars were calculated using the Least significant Difference test at P < 0.05. Multivariate analyses were performed on the mean density of A. lycopersici versus predator mites, insect densities, and densities of the different glandular and non-glandular trichomes of the six tomato cultivars versus A. lycopersici and its predator densities under natural conditions in 2015 using the JMP 7 statistical package (SAS, 2007).
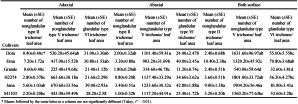
The mean numbers of A. lycopersici mobile stages differed significantly among the six tomato cultivars under controlled conditions (Table 1, F5,48=9.21, P<0.01). Similarly, the mean mite numbers differed significantly among both the abaxial and adaxial surfaces of the six tomato cultivars ( F5,48=7.77, P<0.01, F5,48=9.31, P<0.01), and the mean mite number for Jana was significantly higher than the other cultivars. Although the H2274 cultivar had fewer mites, the difference was not significant compared with the other tomato cultivars.


The A. lycopersici density among the six tomato cultivars varied remarkably throughout the entire growing season under natural conditions (Table 2, F5,409=2.93, P=0.0013). Aculops lycopersici on M1103 had a significantly higher population density, followed by the Etna and Jana cultivars, respectively. Similar to the controlled condition results, mean mite numbers were significantly lower on the Grande and H2274 cultivars than the others. Mean mite numbers significantly differed on both the lower and upper surfaces of these tomato cultivars (Table 2, F5,409=2.50, P=0.03, F5,409=3.59, P=0.0034). The population trends and densities of A. lycopersici on the six tomato cultivars under natural conditions for 2015 are shown in Figure 2A. The mite density increased in early August and peaked in late August. The highest A. lycopersici density was significant on 24 August for Jana and H2274 and on 31 August for the M1103, Dora, Grande and Etna cultivars ( P<0.01). The population density gradually decreased from mid-September to mid-October.


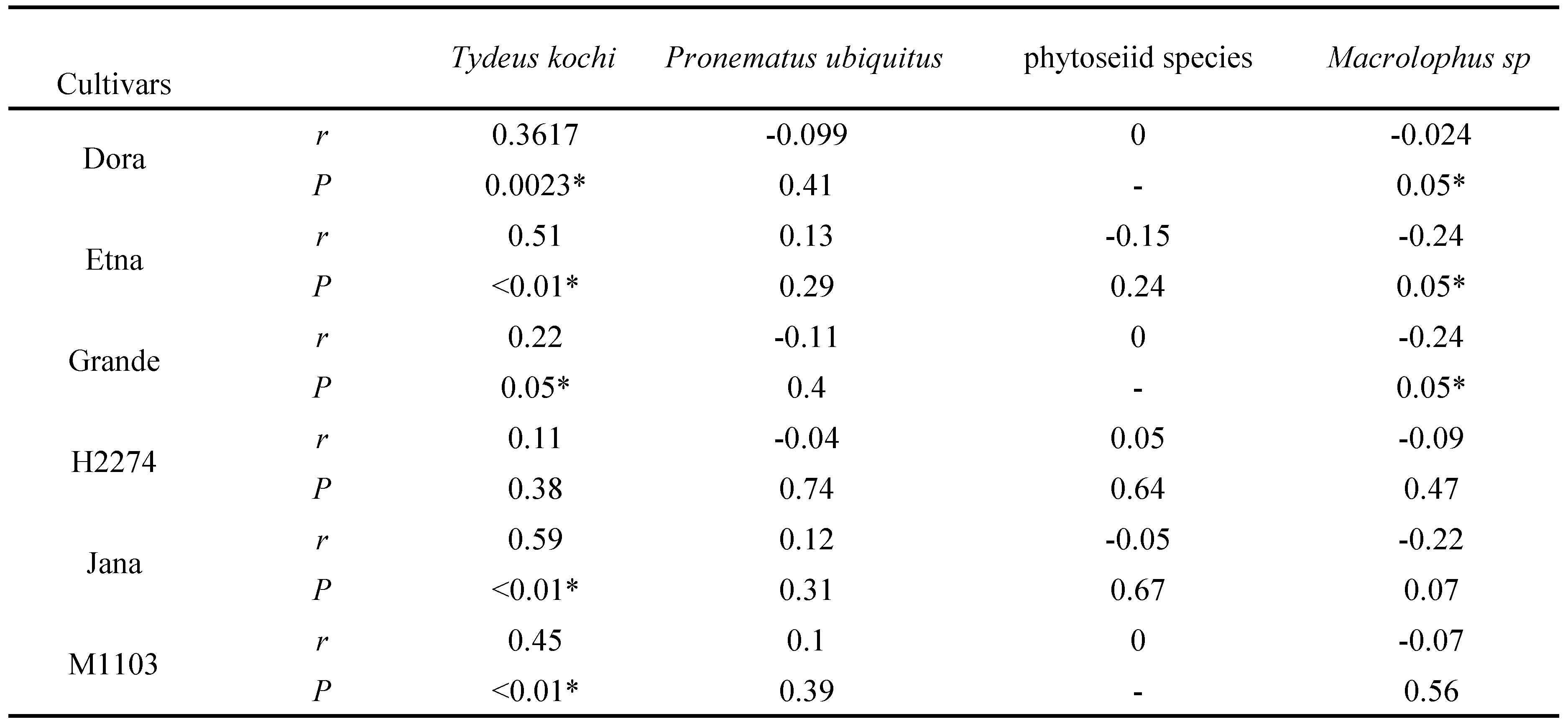
In the present study, 5 predator species belong to Phytoseiidae were obtained from the six tomato cultivars under natural conditions. These species were Neoseiulus barkeri Hughes, Euseius finlandicus (Oudemans), Typhlodromus (Anthoseius) recki Wainstein, Typhlodromus (Typhlodromus) athiasae Porath and Swirski and Phytoseius finitimus Ribaga (Acari: Phytoseiidae). In addition, two species were detected from Tydeidae and Iolinidae, Tydeus kochi Oudemans and Pronematus ubiquitus (McGregor), which have varied feeding habits including predatory.
Tydeus kochi density varied significantly among the six tomato cultivars throughout the entire growing season under natural conditions (Table 3, F5,409=2.28, P=0.046). The highest significant density of T. kochi was found on the H2274 and Grande cultivars. In contrast to the A. lycopersici results, mean mite numbers were significantly lower on the Jana and Dora cultivars than the other cultivars (Table 3). T. kochi population trends and densities on the six tomato cultivars under natural conditions for 2015 are shown in Figure 2B. Similar to the A. lycopersici trend, T. kochi was first observed in late July to early August and peaked significantly twice on 31 August and 20 September ( P<0.01). The T. kochi population trend was positively correlated with that of A. lycopersici on all tomato cultivars (Table 4, r= 0.22 to 0.59, P<0.01 or <0.05). In addition, although the P. ubiquitus densities varied significantly among the six tomato cultivars, they were not significantly correlated with the A. lycopersici population trend (Table 3 and 4, Figure 2C, F5,409=2.79, P=0.017, r=-0.11 to 0.12, P>0.05). However, although phytoseiid were observed between early August and early September, these population trends were not correlated with those of A. lycopersici, and the difference in their population density among the six tomato cultivars was not significant (Tables 3 and 4, Figure 2D, P>0.05, r= -0.15 to 0.05, P>0.05).




In this study, the mite predatory species, Macrolophus sp. (Heteroptera: Miridae) was observed on all tomato cultivars under natural conditions (Table 3 and Figure 2E), but the cultivar did not significantly affect the population density of the predatory insect ( F5,409=0.69, P=0.63). The Macrolophus sp. density was slightly higher on the Grande, Etna and M1103 cultivars. Similar to A. lycopersici, the predator insect was first observed in early August (Figure 2E). In contrast, the highest significant population densities were seen on 16 September and 5 and 19 October ( P<0.01); however, these were not positively correlated with the A. lycopersici population density (Table 4, r=-0.24 to -0.02).

Non-glandular (types II and V) and glandular trichome (type VI) densities on the tomato leaves under natural conditions differed significantly (Table 5, type II F5,24=8.91, P<0.01, type V F5,24=57.55, P<0.01, type VI F5,24=19.13, P<0.01). While trichome density was higher on the abaxial surface for all cultivars, their densities on both surfaces differed significantly among the six tomato cultivars (type II F5,24=9.99, P<0.01, F5,24=5.97, P<0.01, type V F5,24=26.99, P<0.01, F5,24=63.57, P<0.01, type VI F5,24=7.53, P<0.01, F5,24=32.07, P<0.01). The trichome densities of non-glandular type II, and especially type V, on both surfaces were highest on Jana, followed by H2274 and Dora. The lowest trichome densities were detected on Grande (Table 5). Similarly, glandular type VI trichomes were high for Jana and low for Grande (Table 5).



The present study showed that differences in tomato plant cultivars can significantly affect A. lycopersici population densities in both controlled and natural conditions. The mite population density was significantly higher in the Jana cultivar followed by the Etna and M1103 cultivars. However, Grande and H2274 had lower mite population densities. These findings are consistent with other studies that reported differences among tomato cultivars/accessions or wild tomato species infested with A. lycopersici (Kamau et al., 1992; Leite et al., 2000; Kitamura and Kawai 2006; Wang et al., 2008).
Under natural conditions, the A. lycopersici density reached its highest yearly population from mid-August to mid-September, when the mean temperature and relative humidity were 25.6°C and 60.9%, respectively. Consistent with our results, the optimum conditions for A. lycopersici have shown to be 26-29°C and 53-75% relative humidity (Baradan-Anakari and Daneshvar 1992; Kim et al., 2002; Haque and Kawai 2002; Kawai and Haque 2004; Xu et al., 2006). In accordance with our findings, it is reported that the mites first emerge in early August when the mean temperature reaches 26.1-28.3°C and the tomato phenology is in the beginning of the fruit ripening period in north and western Turkey (Hıncal et al. 2002; Yanar et al., 2008). These researchers noted that the mite population and damage gradually increased in mid-August and peaked in early September.
Our results showed significant differences in both glandular and non-glandular trichome densities among the six tomato cultivars. In this study, only non-glandular type II and V and glandular type VI trichomes were observed on all six tomato cultivars, because we did not use a wild cultivar (McDowell et al., 2011). Remarkably, the greatest significant numbers of glandular and non-glandular trichomes were found on the Jana cultivar, followed by Etna, which both had higher mite population densities. Similar to the population density results for A. lycopersici, both trichome densities were much smaller in the Grande cultivar than the other cultivars. In addition, high type V and type VI trichome densities positively affected the population density of A. lycopersici. This effect likely occurred because the mite is smaller than the trichomes (Van Houten et al., 2013b), confirming our hypothesis that trichomes provide excellent shelter for pest mites. Seven species of predatory mites (N. barkeri, E. finlandicus, T. ( A.) recki, T. (T.) athiasae, P. finitimus, P. ubiquitus and T. kochi) obtained in the current study were found to be associated with A. lycoperesici in previous studies (Bayan 1998, Can and Çobanoğlu 2010; Momen and Abdel-Khalek 2008; Çobanoğlu and Kumral 2014; Kumral and Çobanoğlu 2015a,b). Among these species, few phytoseiids were found among all tomato cultivars, but there were no significant differences between the six tomato cultivars with different trichome densities in this study. As mentioned in previous studies, glandular and non-glandular trichomes may affect phytoseiid populations (Fischer and Mourrut-Salesse 2005; Sato et al., 2011; Van Houten et al., 2013a;b). Thus, among glandular trichomes, type VI, is an important defence for tomato plants that entrap arthropods and phytoseiids (Kennedy 2003). The non-glandular trichome density can also limit phytoseiid prey-searching behaviours (Van Houten et al., 2013b). Although it was reported by Van Houten et al. (2013b) that the phytoseiid mite, Amblydromalus limonicus Garman and McGregor suppressed A. lycopersici due to glandular trichome destruction by T. urticae in the early period our results confirmed that the trichomes influenced phytoseiid density under natural conditions (in the presence of T. urticae). In our study, the T. kochi density showed a similar pattern to that of A. lycopersici, and the mite was associated with A. lycopersici populations. Consistent with our results, some tydeid species as an unspecialized group, e.g., T. kochi, Tydeus californicus (Banks), Tydeus caudatus Dugès, Tydeus caryae Kanjani & Ueckermann, appeared to associate with eriophyid members (Rasmy 1971; Khanjani and Mirab-balou 2005; Lorenzon et al., 2009; Çobanoğlu and Kumral 2014). Similarly, Rasmy (1971) reported that T. kochi was a biological control agent of the citrus brown mite, Eutetranychus orientalis (Klein) (Acari: Eriophyidae), in citrus orchards in Egypt. Remarkably, this study found that T. kochi and A. lycopersici densities were smaller and greater, respectively, regarding which cultivars had higher trichome densities. Thus, high type II and type VI trichome densities negatively affected the population density of T. kochi. These findings support the hypothesis that trichome-mediated defences in tomato plants implicated negative tritrophic effects mediated by direct (entrap and kill predators, increasing predation time) and indirect effects (limit development, modify food/prey quality) on natural enemies with trichomes (Farrar and Kennedy 1987; Drukker et al. 1997, Van de Boom et al. 2003; Kennedy 2003; Simmons and Gurr 2005; Koller et al., 2007, Buitenhuis et al. 2015). In addition to tydeid mites, the Iolinidae mite, Pronematus ubiquitus, obtained in the current study, has different feeding habits including predation, phytophagy, fungivory and scavenging. Both tydeid and iolinid populations can persist when eriophyid levels are low because their diets include non-prey foods such as pollen and fungi (Hessein and Perring 1986; Lorenzon et al., 2009). Because of these feeding habits, the mites have never reduced eriophyid densities below the economic threshold as phytoseiids (Gerson et al., 2003). The predator insect, Macrolophus sp., shows a delayed density-dependent numerical response to A. lycopersici density from early September to early October. In contrast to predator mites, the insect density was unaffected by varietal differences, probably due to the larger insect size. However, the non-glandular trichome type V density negatively affected the Macrolophus sp. density. This result was consistent with Nannini and Carboni (2003) who found that Macrolophus caliginosus Wagner (Heteroptera: Miridae) reduced the A. lycopersici density. Regarding tritrophic interactions, Economou et al., (2006) indicated that, except for the time spent grooming, Macrolophus pygmaeus Rambur and Orius niger Wolff nymphs’ probing and moving activities were unaffected by extensive trichome density on the tomato cultivars.
In conclusion, both bitrophic and tritrophic interactions tested under natural conditions showed that the Grande cultivar was a less suitable host for A. lycopersici. Before making a selection from hundreds of tomato cultivars for growing, the cultivar should be examined to assess whether it is unfavourable to A. lycopersici while also being suitable to their predators, such as phytoseiids, tydeids and iolinids. The present study may provide evidence for developing new tomato russet mite-resistant tomato lines.
This study is a part of Elif AYSAN’s MSc thesis, which was completed at the University of Bursa Uludag, Graduate School of Natural and Applied Science, Entomology Department. We thank Professor Edward Albert Ueckermann (ARC-Plant Protection Research Institute, Queenswood, Pretoria, 0121, South Africa) for confirming the mite identification.
Anonymous, 2015. BKU DATABASE. https://bku.tarim.gov.tr/ ![]()
Alba J.M., Montserrat M., Fernández-Mu-oz R. 2009. Resistance to the two-spotted spider mite (Tetranychus urticae) by acylsucroses of wild tomato (Solanum pimpinellifolium) trichomes studied in a recombinant inbred line population. Experimental and Applied Acarology, 47: 35-47. doi:10.1007/s10493-008-9192-4 ![]()
Antonious G.F., Snyder J.C. 2006. Natural products: repellency and toxicity of wild tomato leaf extracts to the two-spotted spider mite, Tetranychus urticae Koch.. Journal of Environmental Science and Health. Part B, Pesticides, Food Contaminants, and Agricultural Wastes, 41: 43-55. doi:10.1080/03601230500234893 ![]()
Aragão C.A., Dantas B.F., Benites F.R.G. 2002. Effect of allelochemicals in tomato leaf trichomes on mite (Tetranychus urticae Koch.) repellency in genotypes with different levels of 2-tridecanone. Acta Botanica Brasilica, 16: 83-88. doi:10.1590/S0102-33062002000100010 ![]()
Baker, E.W. 1970. The genus Tydeus: Subgenera and species groups with descriptions of new species (Acarina: Tydeidae). Annals of the Entomological Society of America, 63(1): 163-177. doi:10.1093/aesa/63.1.163 ![]()
Baradan-Anakari P., Daneshvar H. 1992. Studies on the biology and chemical control of tomato russet mite, Aculops lycopersici (Acari: Eriophyidae), in Varamin. Applied Entomology and Phytopathology, 59(1-2): 25-27.
Bayan A. 1998. Injurious and predatory mites on plants in Lebanon. Arab Journal of Plant Protection, 16(2): 94-97.
Brodeur J., Bouchard A., Turcotte G. 1997. Potential of four species of predatory mites as biological control agents of the tomato russet mite, Aculops lycopersici (Massee) (Eriophyidae). Canadian Entomology, 129(1): 1-6. doi:10.4039/Ent1291-1 ![]()
Buitenhuis R., Murphy G., Shipp L., Scott-Dupree C. 2015. Amblyseius swirskii in greenhouse production systems: a floricultural perspective. Experimental and Applied Acarology, 65 (4): 451-464. doi:10.1007/s10493-014-9869-9 ![]()
Can M., Çobanoğlu S. 2010. Studies on the determination of mite (Acari) species and their hosts of greenhouse vegetables in Kumluca, Antalya. Ziraat Fakültesi Dergisi, 23(2): 87-92.
Celar F., Valič N. 2003. Tomato russet mite (Aculops lycopersici) (Tryon, 1917) (Eriophydae) in Slovenia. Anthology of Papers 6th Slovenian Conference on Plant Protection, Zreče, Slovenian, 489-492.
Channarayappa C., Shivashankar G., Muniyappa V., Frist R.H. 1992. Resistance of Lycopersicon species to Bemisia tabaci, a tomato leaf curl virus vector. Canadian Journal Botany, 70(21): 2184-2192. doi:10.1139/b92-270 ![]()
Chatzivasileiadis E.A., Sabelis M.W. 1998. Variability in susceptibility among cucumber and tomato strains of Tetranychus urticae Koch to 2-tridecanone from tomato trichomes: effects of host plant shift. Experimental and Applied Acarology, 22: 455-466. doi:10.1023/A:1006077521610 ![]()
Çobanoğlu S., Kumral N.A. 2014. The biodiversity and population fluctuation of plant parasitic and benificial mite species (Acari) in tomato fields of Ankara, Bursa and Yalova provinces. Turkish Journal of Entomology, 38(2):197-214.
Drukker B., Janssen A., Ravensberg W., Sabelis M.W. 1997. Improved control capacity of the mite predator Phytoseiulus persimilis (Acari: Phytoseiidae) on tomato. Experimental and Applied Acarology, 21: 507-518. doi:10.1023/B:APPA.0000018885.35044.c6 ![]()
Economou L.P., Lykouressis D.P., Barbetaki A.E. 2006. Time allocation of activities of two heteropteran predators on the leaves of three tomato cultivars with variable glandular trichome density. Environmental Entomology, 35(2): 387-393. doi:10.1603/0046-225X-35.2.387 ![]()
Faraji F., Çobanoğlu S., Çakmak, I. 2011. A checklist and a key for the Phytoseiidae species of Turkey with two new species records (Acari: Mesostigmata). International Journal of Acarology, 37(1): 221-243. doi:10.1080/01647954.2011.558851 ![]()
Farrar R.R. Jr., Kennedy G.G. 1987. 2-Undecanone, a constituent of the glandular trichomes of Lycopersicon hirsutum f. glabratum: effects on Heliothis zea and Manduca sexta growth and survival. Entomologia Experimentalis and Applicata, 43: 17-23. doi:10.1111/j.1570-7458.1987.tb02196.x ![]()
Fischer S., Mourrut-Salesse J. 2005. Tomato Russet Mite in Switzerland (Aculops lycopersici: Acari, Eriophyidae). Revue Suisse de Viticulture, Arboriculture et Horticulture, 37(4): 227-232.
Gerson U., Smiley R.L., Ochoa R. 2003. Mites (Acari) for Pest Control. Blackwell Publishing, Oxford, UK, Pp: 539.
Goldsmith J. 2004. The Tomato Russet Mite: Aculops lycopersici (Massee). Entomology Circular Ministry of Aggriculture and Lands Boulles Research Station.
Gonçalves M.I.F., Maluf W.R., Gomes L.A.A., Barbosa L.V. 1998. Variation of 2-tridecanone level in tomato plant leaflets and resistance to two mite species (Tetranychus sp.). Euphytica, 104: 33-38. doi:10.1023/A:1018611514287 ![]()
Haque M.M., Kawai A. 2002. Population growth of tomato russet mite, Aculops lycopersici (Acari: Eriophyidae) and its injury effect on the growth of tomato plants. Journal of Acarology Society, 11: 1-10.
Hessein N.A., Perring T.M. 1986. Feeding habits of the Tydeidae with evidence of Homeopronematus anconai (Acari: Tydeidae) predation on Aculops lycopersici (Acari: Eriophyidae). International Journal of Acarology, 12(4): 215-221. doi:10.1080/01647958608683467 ![]()
Hıncal P., Yaşarakıncı N., Çınarlı İ. 2002. İzmir ilinde Domates pas akarı (Aculops lycopersici Massee) (Acarina: Eriophyidae)'nın popülasyon seyri, doğal düşmanları ve kimyasal mücadelesi üzerinde araştırmalar. Bitki Koruma Bülteni, 42(1-4): 9-22.
Hoy M.A. 2011. Agricultural Acarology. Introduction to Integrated Mite Management. CRC Press Taylor & Francis Group, New York, 410 pp. doi:10.1201/b10909 ![]()
Kamau A.W., Mueke J.M., Khaemba B.M. 1992. Resistance of tomato varieties to the tomato russet mite, Aculops lycopersici (Massee) (Acarina: Eriophyidae). International Journal of Tropical Insect Science, 13(03): 351-356. doi:10.1017/S1742758400013618 ![]()
Kang J.H. Feng S., Jones A.D., Marksb M.D., Howe G.A. 2010. Distortion of trichome morphology by the hairless mutation of tomato affects leaf surface chemistry. Journal of Experimental Botany, 61: 1053-1064. doi:10.1093/jxb/erp370 ![]()
Kawai A., Haque M.M. 2004. Population dynamics of tomato russet mite, Aculops lycopersici (Massee) and its natural enemy, Homeopronematus anconai (Baker). Japan Agricultural Research Quarterly: 38(3): 161-166. doi:10.6090/jarq.38.161 ![]()
Kennedy G.G. 2003. Tomato, pests, parasitoids, and predators: tritrophic interactions involving the genus Lycopersicon. Annual Review of Entomology, 48: 51-72. doi:10.1146/annurev.ento.48.091801.112733 ![]()
Khalighi M., Dermauw W., Wybouw N., Bajda S., Osakabe M., Tirry L., Van Leeuwen T. 2016. Molecular analysis of cyenopyrafen resistance in the two‐spotted spider mite Tetranychus urticae. Pest Management Science, 72(1): 103-12. doi:10.1002/ps.4071 ![]()
Khanjani M., Mirab-balou M. 2005. Some natural enemies of eriophyid mites from western
Kim D.G., Park D.G., Kim S.H., Park I.S., Choi S.K. 2002. Morphology, biology and chemical control of tomato russet mite, Aculops lycopersici Massee (Acari: Eriophyidae) in Korea. Korean Journal of Applied Entomology, 41(4): 255-261.
Kitamura T., Kawai A. 2006. Difference of susceptibility to damage from tomato russet mite, Aculops lycopersici (Massee) (Acari: Eriophyidae), among varieties within and between species in genus Lycopersicon. Japan Journal of Applied. Entomology and Zoology, 50(1): 57-61.
Koller M., Knapp M., Schausberger, P. 2007. Direct and indirect adverse effects of tomato on the predatory mite Neoseiulus californicus feeding on the spider mite Tetranychus evansi. Entomologia Experimentalis and Applicata, 125: 297-305. doi:10.1111/j.1570-7458.2007.00625.x ![]()
Kumral N.A., Çobanoğlu S., Tiedt L., Ueckermann E. 2014. Domates Pas Akarının Taramalı Elektron Mikroskobuyla Dış Morfolojisi ve Domatesteki Zarar Belirtileri. Türkiye V. Bitki Koruma Kongresi, 3-5 Şubat 2014, Akka Antedon Hotel, Antalya.
Kumral N.A., Çobanoğlu S. 2015a. A reservoir weed for mites: Datura stramonium L.(Solanaceae) in the vicinity of cultivated solanaceous plants in Turkey. International Journal of Acarology, 41(7): 563-573. doi:10.1080/01647954.2015.1084043 ![]()
Kumral N.A., Çobanoğlu S. 2015b. The potential of the nightshade plants (Solanaceae) as reservoir plants for pest and predatory mites. Turkish Journal of Entomology, 39(1): 91-108. doi:10.16970/ted.55042 ![]()
Leite G.L.D., Picanço M., Marquini da Silva F., Casali V.W.D., Galvan T., Cavalcante T.R.M. 2000. Distribution of Aculops lycopersici on Lycopersicon esculentum and Lycopersicon hirsutum canopy and leaves. Agro-Ciencia, 17(2): 259-263.
Lorenzon M., Pozzebon A., Duso C., Palevsky E., Weintraub P.G., Gerson U., Simoni S. 2009. Notes on the phenology and the biology of Tydeus caudatus Dugès (Acari Tydeidae). IOBC/WPRS Bulletin, 50: 45-49.
Maluf W.R., Inoue I.F., Ferreira R.D.P.D., Gomes L.A.A., De Castro E.M., Cardoso M.D.G. 2007. Higher glandular trichome density in tomato leaflets and repellence to spider mites. Pesquisa Agropecuária Brasileira, 42: 1227-1235. doi:10.1590/S0100-204X2007000900003 ![]()
Maluf W.R., Maciel G.M., Gomes L.A.A., Cardoso M.D.G., Gonçalves L.D., da Silva E.C., Knapp M. 2010. Broad-spectrum arthropod resistance in hybrids between high- and low-acylsugar tomato lines. Crop Science 50: 439-450. doi:10.2135/cropsci2009.01.0045 ![]()
Mason P.G., Huber J.T. 2001. Aculops lycopersici (Massee), Tomato Russet Mite (Acari: Eriophyidae): Biological Control Programmes in Canada. Ed.: Mason, P.G., Huber, J.T., Wallingford, UK, pp: 32-33.
McDowell E.T., Kapteyn J., Schmidt A., Li C., Kang J., Descour A., Shi F., Larson M., Schilmiller A., An L., Jones A.D., Pichersky E., Soderlund C.A., Gang D.R. 2011. Comparative functional genomic analysis of Solanum glandular trichome types. Plant Physiology, 155(1): 524-539. doi:10.1104/pp.110.167114 ![]()
Momen F.M., Abdel-Khalek A. 2008. Effect of the tomato rust mite Aculops lycopersici (Acari: Eriophyidae) on the development and reproduction of three predatory phytoseiid mites. International Journal of Tropical Insect Science, 28(01): 53-57. doi:10.1017/S1742758408942594 ![]()
Nannini M., Carboni M.A. 2003. A preliminary investigation on the potential of the mirid bug Macrolophus caliginosus Wagner as biological control agent of the tomato russet mite. Colloque international tomate sous abri, protection intégrée, agriculture biologique, Ed.: Roche, L., Edin, M., Mathieu, V., Laurens, F., Avignon, Paris, France, pp: 185-187.
Ozman-Sullivan S.K., Ocal H. 2005. Sebzelerde Bulunan Eriophiyoid Akarlar. GAP IV. Tarım Kongresi Bildirileri, 21-23 Eylül 2005, Şanlıurfa, Türkiye.
Pocoví, M., Gilardón, E., Gorustovich, M., Olsen, A., Gray, L., Hernández, C., Petrinich, C., Collavino, G. 1998. 2-tridecanona y su asociación con la resistencia a la polilla del tomate (Tuta absoluta Meyrick) ya la arañuela roja (Tetranychus urticae Koch). Revista de la Facultad de Agronomía, 103(2), 165-171.
Rasmy, A.H. 1971. Relation between predaceous and phytophagous mites on Citrus. Journal of Applied Entomology, 67(1-4): 6-9.
Royalty R.N., Perring T.M. 1988. Morphological analysis of damage to tomato leaflets by tomato russet mite (Acari: Eriophyidae). Journal of Economic Entomology, 81(3): 816-820. doi:10.1093/jee/81.3.816 ![]()
SAS 2007. SAS Institute. JMP version 7.0.2 Release Notes. Cary, NC: SAS Institute Print Center; 1-20.
Sato M.M., de Moraes G.J., Haddad M.L., Wekesa V.W. 2011. Effect of trichomes on the predation of Tetranychus urticae (Acari: Tetranychidae) by Phytoseiulus macropilis (Acari: Phytoseiidae) on tomato, and the interference of webbing. Experimental and Applied Acarology: 54(1): 21-32 doi:10.1007/s10493-011-9426-8 ![]()
Schie C.C.N.V., Haring M.A., Schuurink R.C. 2007. Tomato linalool synthase is induced in trichomes by jasmonic acid. Plant Molecular Biology, 64: 251-263. doi:10.1007/s11103-007-9149-8 ![]()
Shipp J.L., Gillespie D.R., Ferguson G.M. 2001. Aculops lycopersici (Massee), tomato russet mite (Acari: Eriophyidae): Biological Control Programmes in Canada. Ed.: Mason P.G., Huber J.T., Wallingford, UK, Pp: 32-33.
Simmons A.T., Gurr G.M. 2005. Trichomes of Lycopersicon species and their hybrids: effects on pests and natural enemies. Agricultural and Forest Entomology, 7: 265-276. doi:10.1111/j.1461-9555.2005.00271.x ![]()
Xu X., Li L., Wang D., Hong X., Wu J., Yuan Y., Xie X 2006. Effect of temperature and relative humidity on development and reproduction of the tomato russet mite, Aculops lycopersici (Massee) (Acarina, Eriophyidae). Acta Entomologica Sinica, 9(5): 816-821.
Ueckermann E.A., Grout, T. 2007. Tydeoid mites (Acari: Tydeidae, Edbakerellidae, Iolinidae) occuring on Citrus in southern Africa. Journal of Natural History, 41(37-40): 2351-2378. doi:10.1080/00222930701589921 ![]()
Van de Boom C.E.M., Van Den Beek T.A., Van Dicke M. 2003. Differences among plant species in acceptance by the spider mite Tetranychus urticae Koch.. Journal of Applied Entomology, 127: 177-183. doi:10.1046/j.1439-0418.2003.00726.x ![]()
Van Houten Y., Knapp M., Hoogerbrugge H., Bolckmans K. 2013a. The potential of Amblyseius swirskii as biocontrol agent for Aculops lycopersici on tomatoes. IOBC/WPRS Bulletin, 93: 51-57.
Van Houten Y.M., Glas J.J., Hoogerbrugge H., Rothe J., Bolckmans K.J. F., Simoni S., Van Arkel J., Alba J.M., Kant M.R., Sabelis M.W. 2013b. Herbivory-associated degradation of tomato trichomes and its impact on biological control of Aculops lycopersici. Experimental and Applied Acarology, 60(2): 127-138. doi:10.1007/s10493-012-9638-6 ![]()
Wang M.Y., Wang D.S., Hong X.Y., Yuan Y.D., Gan J.H. 2008. Relationship between physical and chemical differences of tomato leaf and the resistance to Aculops lycopersici. Chinese Bulletin of Entomology, 45(6): 904-908.
Yanar D., Ecevit O., Kadıoğlu İ. 2008. Tokat Yöresinde domates ekim alanlarında zarar oluşturan Domates pas akarı [Aculops lycopersici (Massee) (Acari: Eriophyidae)]. GaziOsmanPaşa Üniversitesi Ziraat Fakültesi Dergisi, 25(2): 1-5.
Yu S.J. 2008. The toxicology and biochemistry of insecticides. London: CRC Press, 276 pp.



2016-11-21
Date accepted:
2018-01-23
Date published:
2018-09-28
Edited by:
Tixier, Marie-Stéphane

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Aysan, Elif and Kumral, Nabi Alper
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)