



Microarthropods in australian sugarcane soils: A survey with emphasis on the Mesostigmata as potential regulators of nematode populations
Walter, David E.1 and Stirling, G.R.2
1✉ University of the Sunshine Coast, Sippy Downs; Queensland Museum, Brisbane, Australia; Department of Biological Sciences, The University of Alberta, Edmonton, Canada.
2Biological Crop Protection Pty. Ltd., Brisbane, Queensland, Australia.
2018 - Volume: 58 Issue: 3 pages: 673-682
https://doi.org/10.24349/acarologia/20184264Keywords
Abstract
‘Microarthropod’ is a convenient term for describing the minute arthropods that live in soil-litter systems and help regulate many decomposer subsystem processes through their feeding and other behaviours (Moore et al., 1988). Though definitions vary, arthropods more than a few millimetres in length are generally treated as a separate functional group (macroarthropods), although in reality the size distribution is more or less continuous and the smaller size classes interact with much larger animals, particularly in the litter layer. One aspect that links microarthropods worldwide, however, is that they belong to poorly known taxonomic groups such as the mites, springtails, proturans, dipurans, pauropods, symphylans, pseudoscorpions, the tiniest of myriapods, spiders, and insects, especially beetles, ants, parasitoid wasps, and flies. Thus, few areas in the world have the taxonomic infrastructure to deal with microarthropods at the species level. This taxonomic impediment is especially acute in Australia.
Although sugarcane is the most important agricultural crop in tropical and sub-tropical regions of Australia, and a major crop worldwide, the soil microarthropod fauna of cane fields is little studied. Most research has concentrated on the biology and control of relatively large root-feeding pests such as cane grubs and soldier flies, on a few smaller pests such as ground pearls and symphylans, or on arthropod communities at a low level of taxonomic resolution (Allsopp et al., 1993; Sajjad et al., 2012; Shakir and Ahmed, 2015; but see Asif et al., 2016, on Collembola). In this paper we present survey results that show that sugarcane soils, at least those that have not been tilled for several years, contain a diverse community of microarthropods with the potential to improve crop health through their role in cycling nutrients and regulating populations of root pests and pathogens. Initial results from this work have been published previously (Manwaring et al., 2015).
The number of microarthropods extracted from our survey samples was highly variable, raising questions as to the causes of this variability. Previous studies in natural and agricultural ecosystems have shown that the abundance and diversity of microarthropods is influenced by factors such as soil texture, soil carbon levels, depth in the soil profile and the presence of plant residues on the soil surface, and all these factors could have affected our survey results. Consequently, we set up field experiments to look at the impact of mulching, as it is a common practice in the Australian sugar industry and our survey data suggested that the presence of mulch (the residues remaining after harvest and referred to as a ‘trash blanket’ in Australia) may have increased the abundance of microarthropods (see also Manwaring et al. 2018). We also set up a pot experiment to determine whether properties such as the texture of sugarcane soils affected the presence and diversity of mites. Since Mesostigmata seem to be important predators of nematodes (Stirling et al., 2017), we were particularly interested in the impact of mulching and soil properties on this group of mites.
During the period from July to December 2014, 60 mineral soil samples were collected from four major cane-growing regions of Queensland (14 samples from the Herbert; 10 from the Burdekin; 15 from the Mackay region and 21 from farms around Bundaberg, Childers and Hervey Bay). Since tillage is known to be detrimental to microarthropods (Wardle, 1995), samples were, wherever possible, collected from farms where tillage had been minimised and a best-practice farming system had been adopted (Garside et al., 2005). However, third and fourth ratoon crops were always sampled, as this meant that the soil would not have been tilled for at least 3 years. To avoid the negative effects of compaction caused by harvest and haul-out machinery, samples were always collected in the centre of the row and between sugarcane plants. Each sample (about 3 L of soil) was taken with a shovel to a depth of 10 cm from six randomly selected points in a field. Microarthropods were extracted by placing 600 mL of soil on a Tullgren funnel for 5 days (Walter and Krantz, 2009) and the animals were captured in vials containing 70% ethanol. Microarthropods were sorted and counted under a dissecting microscope by feeding guild (see below) and representatives were slide-mounted for further identification when needed.
Because soil microarthropods are extremely diverse, tend to have patchy distributions, and have feeding behaviours that often cross taxonomic boundaries, we sorted our collections into functional groups (guilds) that were based on our preliminary samples (Manwaring et al., 2015) and reflected their importance to the behaviour of interest: nematophagy. Previous research on nematophagy by microarthropods in non-sugarcane soils (reviewed in Walter and Proctor, 2013; Manwaring et al., 2015) documented that this behaviour was pervasive in one order of mites (Acari: Parasitiformes: Mesostigmata). We call these mites the Mesostigmata Guild (i.e. all Mesostigmata collected except the specialised fungivores in the Ameroseiidae). For some analyses we also abstracted a ‘Small Mesostigmata Guild’ (combined totals for all species of Asca, Gamasellodes, Protogamasellus, Multidentorhodacarus, Rhodacarus, and Rhodacarellus). These mites are less than 0.35 mm in length, have divided dorsal shields and narrow bodies (Asca species are at the upper length and width of this group) and are capable of using small pore spaces in soil. The larger Mesostigmata were members of the Ascidae, Ologamasidae, Blattisociidae, Laelapidae, Macrochelidae, Parholaspididae, Pachylaelapidae, and Uropodoidea (see results for identified genera) with inflexible dorsal shields and presumably less access to small pore spaces.
The most diverse and abundant taxa of microarthropods present were oribatid mites (Acari: Acariformes: Oribatida [including Astigmata]) and springtails (Hexapoda: Collembola). As far as is known, most species in these groups feed primarily on microbes and decaying organic matter, but many may consume nematodes when they encounter them (Walter and Proctor, 2013; also see Results). We combined the counts for these two taxa into the Fungivore/Detrivore Guild because in general oribatid mites and springtails share similar feeding behaviours and are thought to have some impact on nematode numbers. The other microarthropods present are predators of arthropods, have more specialised feeding habits or have unknown feeding behaviours. These include proturans, diplurans (Campodeidae, Parajapygidae), assorted insects (thrips, planthoppers, ants, beetles, flies, etc.), millipedes (mostly Polyxenidae), centipedes, small spiders, pseudoscorpions, palpigrades (Eukoeninia sp.), and certain mite groups (mostly Prostigmata [e.g. Eupodidae, Tydeidae, Tarsonemidae, Cryptognathidae, Cunaxidae: Cunaxinae, Bdellidae, Rhagidiidae] and Endeostigmata). We group these as Miscellaneous Arthropods and consider that they are unlikely to be important predators of nematodes in sugarcane soils, although they may provide other ecosystem services.
We tested our assumptions of nematophagy using live extractions (i.e. into vials with a plaster-of-Paris/charcoal base instead of ethanol as per Walter and Krantz, 2009) of microarthropods collected from cane fields in the Bundaberg region, adding water suspensions of a bacterial-feeding nematode cultured on rolled oats and observing the interactions. All observations of nematophagy were recorded. Mesostigmatans observed feeding on nematodes were transferred to small rearing vials with a small paintbrush and nematodes in a water suspension were regularly added to determine if the mites could reproduce and develop to adults on a nematode diet at room temperature. If so, representatives were slide-mounted and identified as far as possible.
To assess the impact of green-cane trash blanketing (i.e. mulching) on the microarthropod community, mulched and non-mulched plots were established at two sites in Bundaberg. The soil at site 1 was a clay loam while site 2 had a lighter-textured sandy clay loam soil. Based on the Australian Soil Classification, these soils were classified as a Red Ferrosol and Yellow Dermosol, respectively. Sugarcane was planted at both sites following soybean and the plant crops were harvested in early September 2015, when the sugarcane was about 13 months old. Two weeks after harvest, the treatments at each site were established by marking out 12 plots 10 m long and 1.85 m wide, raking the trash blanket from half the plots and retaining the crop residues in the mulched plots. Thus, each experiment consisted of two treatments (bare and mulch) x 6 replicates. All plots were 2 m apart and contained one row of sugarcane. Any weeds that grew in the plots (particularly the plots that were not mulched) were either removed periodically by hand or eliminated by the herbicide program used by the grower.
On 10 March 2016, nearly six months after the experiment was set up, samples of mulch and soil were collected to assess populations of nematodes and microarthropods. All samples were collected in the bed and were taken within 15 cm of a sugarcane plant. In the mulched plots, mulch was removed from a 20 x 20 cm sampling zone, and after it was weighed and cut into pieces less than 1 cm long, a 10 g sub-sample from each plot was spread on a nematode extraction tray (Whitehead and Hemming, 1965). After 3 days, nematodes were recovered by sieving twice on a 38 µm-aperture sieve. Microarthropods were recovered from the mulch by placing 15-20 g sub-samples in a Tullgren funnel. Four soil cores 10 cm in diameter were collected to a depth of 5 cm from each of the plots where the mulch was present and from similar positions in the non-mulched plots. The soil in the cores collected from each plot was then mixed gently and nematodes and mites were extracted by placing 230 and 500 g sub-samples of moist soil on trays and funnels, respectively. Soil and mulch samples were also dried at 80°C to determine the dry weight equivalent of each sample. Numbers of microarthropods and nematodes recovered from the sub-samples and the weights of soil and mulch in the processed samples were used to calculate the number of microarthropods and nematodes /kg dry soil and the number of microarthropods and nematodes /10 g dry mulch.
This experiment aimed to determine whether soil properties (in this case soil texture and depth in the soil profile) influenced the microarthropod community when mineral soil is mulched with sugarcane residue. The soil used for the experiment was obtained from six sugarcane fields in Bundaberg that were growing crops ranging in age from third to sixth ratoon and were on two different soil types. Three of the fields had sandy loam soils and three had clay loam (ferrosol) soil. In November 2014, soil was collected from each field at two depths in the soil profile (0-2 cm and 10-15 cm). Equal volumes of soil from each soil type and depth were then mixed together to produce four soils (sandy loam 0-2 cm; sandy loam 10-15 cm; clay loam 0-2 cm; clay loam 10-15 cm). Basic chemical properties such as pH, electrical conductivity, total C and N (Leco method) and labile C (method of Blair et al., 1995) were assessed by Environmental Analysis Laboratory, Lismore, NSW, while microbial activity was measured using the Solvita™ CO2-burst protocol (Franzluebbers, 2016). These analyses showed that the four soils had markedly different properties. Carbon levels and microbial activity were much higher in the clay loam soil than the sandy loam soil and there were similar differences between soils collected from different depths (Table 1).


Four 1.4 L pots were filled with 1 kg dry weight equivalent of each of the soils and pots were planted with pre-germinated single-eye setts of sugarcane (variety Q208). The 16 pots were then placed in a greenhouse and the soil surface was covered with a layer of sugarcane residue (22 g per pot) to simulate the 15.5 t/ha trash blanket that is laid down when a 100t/ha sugarcane crop is harvested (Mitchell and Larsen, 2000). After 8 weeks, soil and roots were removed from the pots and nematodes and microarthropods were extracted by placing 210 g and 400 g of moist soil on trays and funnels, respectively.
Data were analysed by analysis of variance (ANOVA) using Genstat 8.2. The two experiments on the effects of mulching were analysed separately while the pot experiment was analysed as a soil texture x depth factorial. Microarthropod and nematode numbers were transformed (log10 no. animals + 1) prior to analysis. Means ± SE were calculated and regression analyses undertaken using features available in Microsoft Excel.
A total of 60 cane fields were sampled and a trash blanket had been retained as mulch in 50 of these sites while the remaining 10 sites lacked a trash blanket. In total, 7517 microarthropods were extracted and assigned to our three guilds. Numbers per 600 mL sample of mineral soil were highly variable (range of 4-539 in trash-blanketed sites vs 6-185 in sites with no trash blanket), but the animals were almost twice as abundant where the shelter of a trash layer was present (mean = 135.6±16.3 vs 73.9±19.5). The Burdekin sites generally had the lowest numbers of microarthropods, but there were no obvious effects of crop stage, irrigation, soil type, soil moisture or row spacing.
Our results indicate that Queensland sugarcane soils are dominated in numbers by our Fungivore/Detritivore Guild composed of oribatid mites (Acariformes: Oribatida) and springtails (Hexapoda: Collembola) (range 2-415 per sample, mean = 77.1±9.9). The oribatid mites typically were representatives of the Galumnidae, Oripodoidea (especially Scheloribatidae, Oribatulidae), Oppioidea, Nothridae, Epilohmanniidae, Lohmanniidae, Tectocepheidae, and Microzetidae. Other oribatid families were rare or sporadic, but a member of the Acaridae (Rhizoglyphus robini Claparéde) was occasionally abundant. Collembola were represented by members of all of the superfamilies (Poduroidea, Entomobryoidea, Neeloidea, Sminthuroidea). The rapid attraction to and consumption of vermiform nematodes was observed repeatedly for members of the oribatid mite families Galumnidae (Galumna sp., Pergalumna sp.) and Scheloribatidae (Scheloribates sp.) in the live extractions. One observation of nematophagy by an unidentified isotomid springtail was observed. Therefore, it seems likely that these animals function in cane soils as opportunistic nematophages, as they are known to do in other soils (see Walter and Proctor, 2013, for a recent review of nematophagy in mites).
Members of the Mesostigmata, the order of mostly predatory mites with numerous known nematophages, were much less abundant (range 1-80 per sample, mean = 14.1±1.9), but diverse (approximately three dozen species). These included members of genera with species known to feed on nematodes, e.g. Rhodacarus, Rhodacarellus (Rhodacaridae); Gamasellodes, Protogamasellus, Asca, Antennoseius (Ascidae); Lasioseius, Cheiroseius (Blattisociidae); Gaeolaelaps, Cosmolaelaps (Laelapidae); Holaspulus (Parholaspididae); Zygoseius (Pachylaelapidae); Antennolaelaps, Athiasella, Gamasiphis (Ologamasidae) and also unidentified Uropodina (some species are nematophagous, but most have unknown feeding habits). Nematophagy was observed in 15 mesostigmatan species from the Bundaberg collections and reproduction on a diet of nematodes was obtained for 11 of those species: Antennolaelaps sp., Asca garmani Hurlbutt, Asca major Womersley, Cheiroseius sp., Cosmolaelaps sp., Gaeolaelaps sp., Gamasellodes bicolor (Berlese), Gamasiphis sp., Holaspulus tenuipes Berlese, Protogamasellus mica (Athias-Henriot) and Protogamasellus sigillophorus Mineiro, Lindquist and De Moraes. The latter (misidentified as Protogamasellopsis sp. in Manwaring et al. 2015) was described from a cornfield in Brazil (Mineiro et al. 2009) and is reported from Australia for the first time. Although many could not be assigned to a described species, those that were included both semi-cosmopolitan mites such as A. garmani, Gaeolaelaps aculeifer (Canestrini), Gamasellodes bicolor, H. tenuipes, and P. mica; and apparently Australasian endemics such as A. major, Gamasellodes adrianae Walter, Holostaspella moderata Berlese, Gaeolaelaps queenslandicus (Womersley), and Zygoseius sarcinulus Halliday.
Numbers in the Miscellaneous Arthropods group ranged from 2-349 individuals per sample, but most samples contained less than 50 such microarthropods, with the few larger numbers due to sampling at the margins of an ant colony or the presence of large numbers of scale insect crawlers. Predators of arthropods, both mites (e.g. Bdellidae, Cunaxidae, Rhagidiidae, Stigmaeidae) and other arthropods (e.g. diplurans such as Parajapyx sp. and unidentified Campodeidae, small spiders and centipedes) were common. Symphylans, some of which are root pests, were rare. Isopods, small millipedes, fly larvae, psocopterans and a variety of other small insects also were collected. Additionally, microarthropods thought to be fungivores, but not known to eat nematodes (e.g. members of the Ameroseiidae, Eupodidae, Nanorchestidae, Terpnacaridae, Tarsonemidae, Pygmephoridae [all Acari]; Ptilliidae [Coleoptera], Acerentomidae [Protura], Pauropodidae [Pauropoda]) were present in small numbers. With one exception, an unidentified cunaxid mite in the subfamily Cunaxoidinae (see Walter and Kaplan 1991), no members in these groups were observed feeding on nematodes in the live extractions.
Dominance by oribatid mites and springtails is generally associated with a fungus-based decomposer system and should be strongly influenced by the amount of organic matter in the soil and retention of sufficient water to maintain fungal growth (Coleman and Crossley, 2004). Both of these parameters should be enhanced by the retention of a trash blanket. This hypothesis is supported by our survey results, as numbers of Fungivore/Detritivores were on average 55% higher under a trash blanket than in the absence of a trash blanket (81.9±11.5 vs 52.9±14.4 animals per sample). The effect of the trash blanket on the Mesostigmata was even more dramatic, with numbers being 206% higher under a trash blanket (15.5±2.1 vs 7.5±2.4). Farming system also seemed to have a strong influence on Mesostigmata with fields under best-practice management averaging 18.6±3.4 Mesostigmatans compared with 10.5±1.7 in fields using other farming systems. Total microarthropods were also much higher in the former system (150.6±23.6 vs 104.5±16.6).
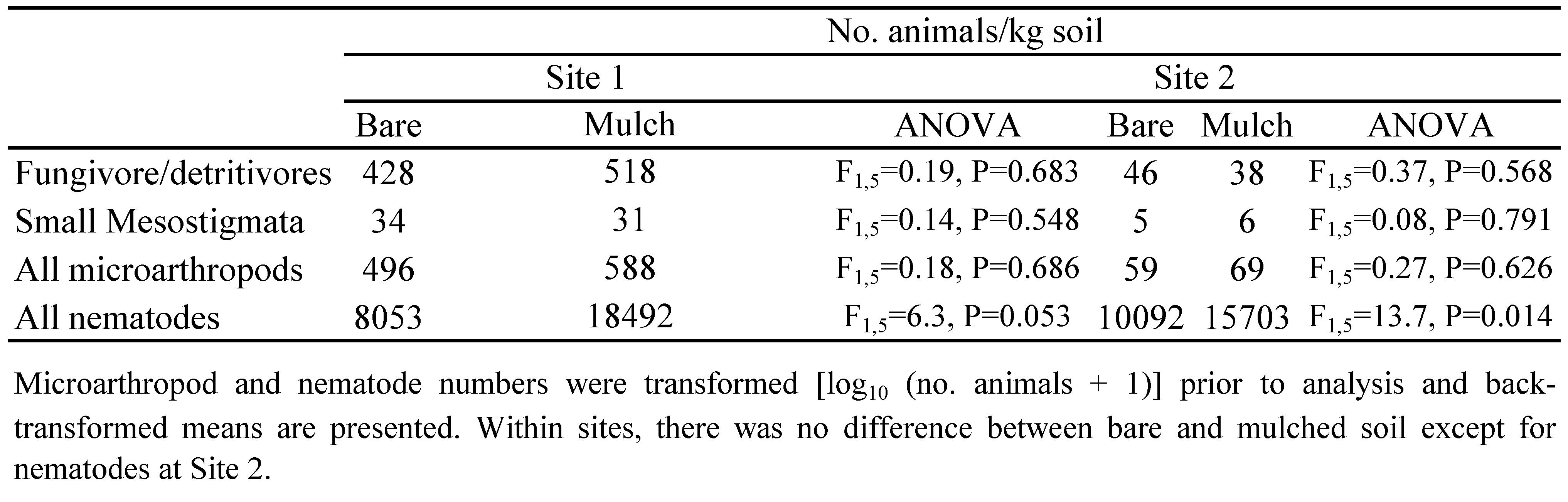
Although the two study sites were less than 10 km apart, there were many more microarthropods at site 1 than site 2 (Table 2). However, analyses of the data from each site indicated that mulching did not significantly affect any component of the microarthropod community, as numbers in mulched and bare plots did not differ significantly. There were more nematodes in the soil under the mulch than the bare soil (Table 2), with statistical analyses indicating that the mulch effect was significant at site 2 and almost significant at site 1. The reason for this was that there were more free-living nematodes (predominantly bacterivores and fungivores) under the mulch. However, there was no relationship between the nematode population and numbers of their microarthropod predators, as numbers of mites in the Small Mesostigmata guild were similar in mulched and bare soil.

The mulch at both sites contained microarthropods but as with the soil, total numbers were much higher at site 1 than site 2 (1123±223 vs 273±30/10 g mulch, respectively). Large numbers of free-living nematodes were also recovered from the mulch at both sites (an average of 8,899 and 12,947 nematodes/10 g mulch at sites 1 and 2 or the equivalent of 222,000 and 324,000 nematodes/m2, respectively). About 10% of the microarthropods in the mulch were nematophagous but the composition of the nematophagous community in the mulch differed from that in soil. The Cunaxoidinae (small, predatory prostigmatid mites) were relatively common in the mulch at both sites (comprising 33 and 32% of the nematophagous community at sites 1 and 2, respectively) whereas they comprised only 12% of the nematophagous community in soil at site 1 and were not recovered from the soil at site 2.
Analyses of total microarthropod numbers showed that there were significant soil texture (2-way ANOVA, F1,9 = 25.6, P < 0.001) and depth effects (F1,9 = 10.8, P = 0.009). The clay loam soil supported many more microarthropods than the sandy loam soil and there were significantly more microarthropods in the topsoil than in soil collected from a depth of 10-15 cm (Table 3). Numbers in the Small Mesostigmata did not differ significantly between soils (F1,9 = 1.17, P = 0.308) but there was a significant depth effect (F1,9 = 55, P< 0.001), with these mites being far more numerous in the topsoil (Table 3). For neither total microarthropods (F1,9 = 0, P = 0.948 nor small Mesostigmata (F1,9 = 4.5, P = 0.062) was the interaction between soil texture and depth significant.


When the nematode data were analyzed the soil texture effect was not significant (F1,9 = 2.75, P = 0.132) but there was a significant depth effect (F1,9 = 20.9, P = 0.001) and a significant soil texture x depth interaction (F1,9 = 9.97, P = 0.012). The most obvious effect was the low number of nematodes in pots filled with the sandy loam soil collected from a depth of 10-15 cm (Table 3). Regression analysis using log-transformed data showed that numbers of mesostigmatans increased as the nematode population increased and that there was a significant but relatively poor relationship between these parameters (P = 0.029; R2 = 0.295).
The microarthropod communities in Queensland sugarcane soils tend to be dominated by the Fungivore/Detritivore Guild (oribatid mites and springtails). These tiny arthropods feed primarily on decomposing plant matter and its associated fungi and other microbes, but many species are known to opportunistically prey on nematodes. Little is known about these animals in sugarcane soils, but only a restricted set of families of oribatid mites are present compared to those found in Australian forest soils (Walter, personal observation). These mites are abundant in well-structured sugarcane soils with mulch layers, and as well as contributing to soil health through their effects on decomposition and nutrient cycling, they may sometimes use nematodes as a food source and possibly act as vectors of microbial nematode antagonists (see Walter and Proctor, 2013).
In contrast, mesostigmatans actively prey on nematodes. Many species require nematodes in their diets for reproduction and Australian species have been shown to exhibit a high degree of nematophagy (Beaulieu and Walter, 2007). In cane soils we found about three-dozen species, many in genera known to have nematophagous species, and observed 15 species feeding on nematodes, including 11 species that were reared successfully with only nematode prey. Many of these are general predators that can subsist on alternative prey (e.g. springtails, mites, small insects), but show improved development and increased reproduction when nematodes are eaten (Walter and Ikonen, 1989). As with the oribatid mites, only a subset of the mesostigmatan fauna was found in cane soils and many of the genera and species that we did find are common in other agricultural systems (see Walter and Kaplan, 1990a, b). Some of these mites are voracious nematophages and if large populations are present, they are likely to contribute to the development of nematode-suppressive soils.
In general, results from the 60 sites surveyed showed that the abundance of all microarthropod groups was highly variable. This is to be expected in a complex system where the soil biology is influenced by many factors. Although no obvious effects of crop stage, irrigation, soil type, soil moisture or row spacing were observed, sites where the trash blanket was retained after harvest and those where the farming system included crop rotation, minimum tillage and controlled traffic showed strong positive effects on the abundance of microarthropods. These effects were especially noticeable with the Mesostigmata, which are thought to be the most important microarthropod predators of nematodes.
Although our field experiments were set up to examine the effect of mulching on microarthropods, the results also showed that numbers of microarthropods were much higher in the clay loam (ferrosol) soil than the sandy clay loam. Ferrosols have much higher soil C levels than lighter–textured soils (see Table 1) and as soil organic matter is the energy source that fuels the soil food web, such a result was expected. However, the textural properties of the clay loam soil probably also contributed, as its polyhedral or blocky structure (McKenzie et al., 1999) meant that habitable pore space was available for microarthropods. Results from our pot experiment also showed that more microarthropods were recovered from the clay loam soil than a much lighter-textured sandy loam, again emphasising the fact that a soil’s textural properties and carbon levels influence microarthropod abundance (Vreeken-Buijs et al., 1998; Larsen et al, 2004; Shakir and Ahmed, 2015).
Since our survey results showed that cane fields with mulch layers tended to have higher populations of microarthropods in mineral soil than fields where the mulch had been removed, and this was consistent with the findings of Manwaring et al. (2018), we assumed that mulching was providing a soil moisture and temperature environment that favoured these animals. Thus, we were surprised that there was no significant difference in the number of microarthropods recovered from mulched and bare soils in our field experiments. However, total numbers of microarthropods at the two sites were 44 and 34% higher in plots that were mulched, suggesting that a mulch effect may have been obtained if more replicates had been included in the experiment or the treatments had been maintained for longer.
One clear result from the two field experiments was that large numbers of microarthropods are able to live in the mulch layer. Since free-living nematodes are also found in high numbers in the mulch, it is not surprising that nematophagous microarthropods are relatively common. However, one interesting observation was that the Cunaxoidinae were much more important component of nematophagous community in the mulch than the soil. This group of mites are comparable in body length to the Small Mesostigmata but have longer legs and are fast moving. We can only speculate as to why they mainly occur in mulch rather than soil but it is possible that they move more easily in the mulch layer due to its greater porosity.
Given the farm to farm variability in microarthropod numbers that we encountered in our survey, we believe that research should be undertaken to understand the main causes of this variability. Soil type and environmental conditions (particularly rainfall and temperature) will certainly be important causal factors but from a sugar industry perspective, it is also important to know whether the practices used to grow the crop are contributing. Our results suggest that microarthropod numbers are higher in soils farmed using best-practice systems but as we surveyed only a limited number of sites, future research should focus on confirming that observation.
With regard to nematophagous mites and their role in regulating populations of plant-parasitic nematodes, it was encouraging to find a diverse range of these animals in sugarcane soils. However, if we are to determine whether they are helping to reduce populations of key nematode pests more work will be required. Since many potential food sources are available in soil, we need to understand the food preferences of individual species and determine whether plant-parasitic nematodes are an important component of their diet in the field. The other key question is whether a particular predator is able to access pest nematodes such as Meloidogyne and Pratylenchus, as they are commonly found at depths of 5-30 cm rather than in the surface soil where most microarthropods are located (Stirling, 2017). This means that researchers must focus on species that live in deeper layers of the soil profile; determine whether parameters such air-filled porosity, bulk density, and body size or shape influence their distribution; and identify the main pathways used by nematophagous mites to access their prey (e.g. the channels that remain when roots decompose or the macropores made by earthworms and other ecosystem engineers).
Sugar Research Australia provided financial support for this study (project 2014/004). We thank Dr Helen Nahrung for help with the statistical analysis.
Allsopp P.G., Chandler K.J., Samson P.R., Story P.G. 1993. Pests of Australian sugarcane. Bureau of Sugar Experiment Stations: Brisbane.
Asif M.U., Ahmed S., Khan R.R., Atiq M. 2016. Relationship of Collembola population with different abiotic factors in an agricultural ecosystem of Faisalabad, Punjab, Pakistan. Pak. J. Agri. Sci., 53: 201-208. doi:10.21162/PAKJAS/16.4179 ![]()
Beaulieu F., Walter D.E. 2007. Predation in suspended and forest floor soils: observations on Australian mesostigmatic mites. Acarologia, 47: 43-54.
Blair G.J., Lefroy R.D.B., Lisle L. 1995. Soil carbon fractions based on their degree of oxidation, and the development of a carbon management index for agricultural systems. Aust. J. Agric. Res., 46: 1459-1466. doi:10.1071/AR9951459 ![]()
Coleman D., Crossley Jr D.A. 2004. Fundamentals of Soil Ecology 2nd Edition. Academic Press: USA. Pp. 408 ISBN: 9780121797263
Franzluebbers A.J. 2016. Should soil testing services measure soil biological activity? Agric. Environ. Lett., 1: 150009. doi:10.2134/ael2015.11.0009 ![]()
Garside A.L., Bell M.J., Robotham B.G., Magarey R.C., Stirling G.R. 2005. Managing yield decline in sugarcane cropping systems. Int. Sugar J., 107: 16-26.
Larsen T., Schjønning P., Axelsen J. 2004. The impact of soil compaction on euedaphic Collembola. App. Soil Ecol. 26: 273-281.
Manwaring M., Walter D., Stirling G.R. 2015. Microarthropods as predators of nematode pests in sugarcane soils: a review and preliminary studies. Proc. Aust. Soc. Sugar Cane Technol., 37: 212-217.
Manwaring M., Wallace H.M., Weaver H.J. 2018. Effects of a mulch layer on the assemblage and abundance of mesostigmatan mites and other arthropods in the soil of a sugarcane agroecosystem in Australia. Exp. Appl. Acarol. 74: 291-300. doi:10.1007/s10493-018-0227-1 ![]()
McKenzie N., Isbell R.F., Brown K., Jacquier D. 1999. Major soils used for agriculture in Australia. In: Peverill K.I., Sparrow L.A. and Reuter D.J. (Eds). Soil Analysis and Interpretation Manual. CSIRO Publishing, Collingwood, p. 71-94.
Mitchell R.D.J., Larsen P.J. 2000. A simple method for estimating the return of nutrients in sugarcane trash. Proc. Aust. Soc. Sugar Cane Technol., 22: 212-216.
Mineiro J.L. De C., Lindquist E.E., De Moraes G.J. 2009. Edaphic ascid mites (Acari: Mesostigmata: Ascidae) from the state of São Paulo, Brazil, with description of five new species. Zootaxa, 2024: 1-32.
Moore J.C., Walter D.E., Hunt H.W. 1988. Arthropod regulation of micro- and mesobiota in belowground detrital foodwebs. Ann. Rev. Entomol., 33: 419-439. doi:10.1146/annurev.en.33.010188.002223 ![]()
Sajjad A., Ahmad F., Makhdoom A.H., Imran A. 2012. Does trash burning harm arthropod biodiversity in sugarcane? Int. J. Agric. Biol. 14:1021-1023.
Shakir M.M., Ahmed S. 2015. Seasonal abundance of soil arthropods in relation to meteorological and edaphic factors in the agroecosystems of Faisalabad, Punjab, Pakistan. Int. J. Biometeorol. 59: 605-616. doi:10.1007/s00484-014-0874-9 ![]()
Stirling G.R. 2017. Soil carbon, root health and nematode pests in sugarcane soils. 1. Root and soil health and its relationship to soil carbon levels. Proc. Aust. Soc. Sugar Cane Technol., 39: 155-165.
Stirling G.R., Stirling A.M., Walter D.E. 2017. The mesostigmatid mite Protogamasellus mica, an effective predator of free-living and plant-parasitic nematodes. J. Nematol., 49:327-333. doi:10.21307/jofnem-2017-080 ![]()
Vreeken-Buijs M.J., Hassink J., Brussaard L. 1998. Relationships of soil microarthropod biomass with organic matter and pore size distribution in soils under different land use. Soil Biol. Biochem. 30: 97-106.
Wardle D.A. 1995. Impacts of disturbance on detritus food webs in agro-ecosystems of contrasting tillage and weed management practices. Adv. Ecol. Res., 26:105-183. doi:10.1016/S0065-2504(08)60065-3 ![]()
Walter D.E., Ikonen E.K. 1989. Species, guilds and functional groups: taxonomy and behavior in nematophagous arthropods. J. Nematol., 21: 315-327.
Walter D.E., Kaplan D.T. 1990a. Antagonists of plant-parasitic nematode pests of citrus in Florida. J. Nematol., 22: 567-573.
Walter D.E., Kaplan D.T. 1990b. A guild of thelytokous mites associated with citrus roots in Florida. Environ. Entomol., 19: 1338-1343.
Walter D.E., Kaplan D.T. 1991. Observations on Coleoscirus simplex (Acarina: Prostigmata), a predatory mite that colonizes greenhouse cultures of rootknot nematode (Meloidogyne spp.), and a review of feeding behavior in the Cunaxidae. Exp. Appl. Acarol., 12: 47-59. doi:10.1007/BF01204399 ![]()
Walter D.E., Krantz G.W. 2009. Collecting, rearing, and preparing specimens. In: Krantz G.W. and Walter D.E. (Eds). A Manual of Acarology 3rd Edition. Texas Tech University Press. Lubbock. p. 83-96.
Walter D.E., Proctor H. 2013. Mites: Ecology, Evolution & Behaviour: Life at a Microscale 2nd Edition. Springer: Netherlands. pp. 494. doi:10.1007/978-94-007-7164-2 ![]()
Whitehead A.G., Hemming J.R. 1965. A comparison of some quantitative methods of extracting small vermiform nematodes from soil. Ann. Appl. Biol., 55: 25-38.



2017-07-05
Date accepted:
2018-04-25
Date published:
2018-07-24
Edited by:
Bonato, Olivier

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Walter, David E. and Stirling, G.R.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)