



Bisexual and oedipal reproduction of Macrocheles muscaedomesticae (Acari, Macrochelidae) feeding on Musca domestica (Diptera, Muscidae) eggs
Farahi, Sara 1 ; Shishehbor, Parviz 2 and Nemati, Alireza 3
1✉ Department of Plant Protection, College of Agriculture, Shahid Chamran University of Ahvaz, Ahvaz, Iran.
2Department of Plant Protection, College of Agriculture, Shahid Chamran University of Ahvaz, Ahvaz, Iran.
3Department of Plant Protection, College of Agriculture, Shahrekord University, Shahrekord, Iran.
2018 - Volume: 58 Issue: 2 pages: 430-441
https://doi.org/10.24349/acarologia/20184251Keywords
Abstract
House flies, Musca domestica L., face flies, Musca autumnalis De Geer, and stable flies, Stomoxys calcitrans L. are the primary dipterous pest species occurring in sheep, cattle, buffalo and poultry farms in Iran (Khoobdel and Davari, 2011). All use animal manure as the substrate for their eggs. The eggs hatch in the manure with the larvae developing and pupating within or under the manure. Current control of these pests still relies mainly on insecticides, although these species have developed resistance to most available insecticides due to their overuse (Keiding, 1999; Sharififard and Safdari, 2013). Increasing awareness in the public and among farmers has encouraged researchers to look for alternative methods of managing these pest flies. The predatory mite, Macrocheles muscaedomesticae (Scopoli) (Acari: Macrochelidae), which is the best known and most abundant species found in cattle and poultry manure in Iran (Fathipour, 1994; Kazemi and Rajaei, 2013), preys on, and is disseminated by many filth flies. M. muscaedomesticae lives in the outermost layers of manure heaps, the region in which flies usually oviposit. This predator feeds mainly on fly eggs (which it prefers) and on first instar larvae (Gerson et al., 2003). The possibility of mass production of M. muscaedomesticae as a candidate biological control agent was evaluated by Filipponi (1964) who verified that Macrochelidae held promise as biological control agents of pest flies. He pointed out that mass production of macrochelids needed elucidation of optimum ecological conditions for each potential mite species, and that the selection of particularly prolific strains that would increase production levels needed to be determined (Filipponi, 1964). Currently, the soil-dwelling predatory mite, Macrocheles robustulus (Berlese) has been commercially available since 2010 for control of thrips and soil inhabiting fly larvae such as Sciaridae (Koppert, 2010).
Although the life history traits and life table parameters of M. muscaedomasticae have been previously studied (e.g. Pereira & de Castro, 1945; Axtell, 1961, 1963; Wade & Rodriguez, 1961; Filipponi, 1964; Cicolani, 1979; Abo-Taka et al., 2014), it has since been shown that different strains (also termed races or ecotypes) of acarine species used as biocontrol agents that originate from different regions, differ in biological attributes that can potentially effect their success rate. For instance, an Israeli strain of Phytoseiulus persimilis Athias-Henriot (Acarina: Phytoseiidae) was more tolerant to low humidity than a California strain (Perring and Lackey, 1989), a trait that affects field survival. Galazzi and Nicoli (1996) compared the life histories of populations of P. persimilis that originated from different regions of Italy, and noted that some lived significantly longer and had higher fecundity than others. Use of pesticide resistant strains of phytoseiids has usually resulted in improved pest control (Gerson et al., 2003). In addition, under natural conditions, especially earlier in the year, virgin female mites may become isolated and reproduce parthenogenetically. Unmated females produce only male offspring; later on, the same mothers can mate with their sons to produce females (Oedipal reproduction) (Ho, 1985; Adamson and Ludwig, 1993). To help understand the complex reproductive biology of M. muscaedomesticae (Iranian strains), we conducted two separate life table experiments in the present study to quantitatively analyze bisexual and oedipal reproduction strategies in M. muscaedomesticae, using the age-stage, two-sex life table which, unlike traditional life tables, includes male individuals and the stage differentiation (Chi and Liu, 1985; Chi, 1988).
The biology and demographic parameters of M. muscaedomesticae were investigated in a growth chamber set at 28 ± 1 °C, a photoperiod of 14:10 (L: D) h and 65 ± 5% relative humidity (RH). Because most of the previous studies related to the life history of M. muscadomesticae were conducted at 28°C (e.g. Cicolani, 1979; Abo-Taka et al., 2014), this temperature was selected in order to maintain similarity in experimental conditions. Specimens of Mu. domestica adults were originally field collected on the campus of Shahid Chamran University of Ahvaz (31°17′59′′N, 48°39′39′′E) and kept in rearing cages (30×30×40 cm). Adult flies were provided with water, powdered milk and sugar. Larvae were reared on a mixture of alfalfa, date extract, wheat bran and water (Sharififard et al., 2011). Housefly eggs were collected daily from larval medium in rearing cages using a fine brush and used as food for mites.
M. muscadomesticae adults were collected from cow manure from the dairy farm on the campus of Shahid Chamran University of Ahvaz and then extracted by Berlese funnel to establish a colony. Stock colonies of mites were maintained in covered, clear, polystyrene storage containers (6 cm dia. × 3.5 cm h.), with ca. 15 cc of sterilized cow manure added. The life table study of M. muscaedomesticae was conducted in plastic containers of 1×1×1 cm rearing cells filled with sterilized cow manure.
Before introducing the mites, water was added to the manure and the surface of the substrate tamped down to facilitate observation of the different mite stages within the rearing cells. In order to maintain the humidity in the rearing cells, water drops were added daily as necessary. Pairs of M. muscaedomesticae were randomly selected from the mite colony and placed in containers. Monitorings were conducted hourly until egg laying occurred, after which adults were removed, and each egg was placed into an individually numbered cell. “Tanglefoot” was applied to the rim to prevent escape by the mites.
In total, 64 eggs were used at the start of the life table experiment. The development time of immature stages was recorded hourly. When the adults eclosed, virgin male and female mites were paired and kept together in rearing containers. Adult survival, longevity, and fecundity data were recorded daily until the death of all individuals. If one member of a pair died, another adult of the same sex and same age was recruited from the mass-rearing colony and paired with the surviving individual. Data relevant to the recruited individual were not included in the data analysis. The eggs laid by each female mite at different ages were kept separately under the aforementioned conditions until reaching adulthood to determine the sex of the offspring.
Eggs used for the study were collected as in the bisexual life table study above. Virgin females were isolated in individual rearing cells after reached the adult stage. Data relevant to development time, survival, and fecundity were analyzed for the 64 females used in the study. New eggs laid by virgin females were moved to separate cells for daily monitoring until mites reached adulthood. The first male offspring developing to the adult stage was retransferred to its mother’s rearing cell to mate with her. Daily survival and fecundity were registered until the death of the mother. Dead male mites were replaced with another adult male from the progeny of the same female. Each egg laid by females after oedipal mating was reared separately until adulthood to record the sex of the progeny.
The life history data of M. muscaedomesticae were analyzed using the age-stage, two-sex life table theory (Chi and Liu, 1985; Chi, 1988) and the computer program TWOSEX-MSChart (Chi, 2017). The survival rate (Sxj) (x = age, j = stage), was calculated as the probability that a newly laid egg would survive to age x and stage j. To take both sexes into consideration, the age-specific survival rate (lx ) was then calculated as:
lx = um_j=1^m Sxj
where m is the number of stages. The age-specific fecundity (mx ) was calculated as:
mx = racum_j=1^m Sxj fxj um_j=1^m Sxj
the population parameters, the intrinsic rate of increase (rm ), the finite rate of increase (λ), the net reproductive rate (R0 ), and the mean generation time (T), were estimated in sequence. The net reproductive rate (R0 ) was calculated as follows:
R0 = um_x=0^nfty lx mx
The intrinsic rate of increase was estimated by using the iterative bisection method from the Euler-Lotka formula:
um_x=0^nfty e-r(extitx+1)lx mx = 1
with age indexed from 0 (Goodman, 1982). The mean generation time, which is defined as the period of time needed by a population to increase to R0 -fold of its size at the stable age-stage distribution, i.e., erT = R0 or λT = R0 , was calculated as T= (ln R0 )/rm .
The reproductive value (vxj ) is considered as the expectation of future offspring of individuals of age x and stage j (Fisher, 1930). Both the age-stage-specific reproduction value (vxj ) and age-stage life expectancy (exj ) were calculated using the program TWOSEX-MSChart (Chi, 2017). The standard errors were estimated by using the bootstrap method (Efron and Tibshirani, 1993; Huang and Chi, 2012; Akköprü et al., 2015), with 100,000 bootstraps to obtain stable estimates of standard errors (Akca et al., 2015). The paired bootstrap test was used to compare differences (Efron and Tibshirani, 1993).
The mean developmental times of different stages are presented in Table 1. Developmental times of all preadult stages, except the egg stage, were significantly different between males and females in bisexual reproduction (Table 1).


The total preadult duration was significantly longer in females than in males in the bisexual cohort ( P < 0.0001). There was no significant difference between total preadult duration of bisexual and oedipal females (P = 0.292).
The mortality of juvenile mites ranged from 1% in oedipal reproduction to 9% in bisexual reproduction (Table 2). Most of the mortality occurred during the protonymph stage of M. muscaedomesticae. There were no significant differences between total preadult mortality in bisexual and oedipal reproducting cohorts (P = 0.80). The sex ratio in bisexual reproduction was 42% (female / total offspring).


Both the adult preoviposition period (APOP) (i.e., the period from adult eclosion to first egg laid) and total preoviposition period (TPOP) (i.e., the period from egg to first egg laid) required for producing female and male offspring were significantly longer in oedipal reproduction than in bisexual reproduction (P < 0.0001) (Table 3). It was calculated for males and females by only including male or female offspring separately in the analysis. Neither the APOP ( P = 0.33301) nor the TPOP (P = 0.33239) was significantly different for producing female over male offspring in the bisexual cohort. As would be expected, both the APOP and TPOP were significantly much longer for producing female than male offspring in the oedipal treatment (P < 0.0001).
The oviposition periods for producing male offspring were significantly longer than those for producing females in both the oedipal (P < 0.0001) and bisexual cohort (P = 0.01683). The oviposition periods for producing male and female offspring in the oedipal cohort were significantly shorter than those in the bisexual group (P < 0.0001) (Table 3). Oedipal female M. muscaedomesticae, on the average, reproduced parthenogenetically during the first 5.62 days of their oviposition period; but produced bisexually during the remainder of their oviposition time following oedipal mating.


The age-stage-specific survival rate (Sxj ) represents the probability that an egg can survive to age x and stage j (Figure 1). Significant overlaps were observed between stages in both of the reproductive cohorts of M. muscaedomesticae due to variable developmental rates occurring among individuals. The ability to observe the beginning and ending of subsequent stages (i.e., protonymph to deutonymph stages) in the survival curve for each stage is an advantage of using the age-stage, two-sex life table.

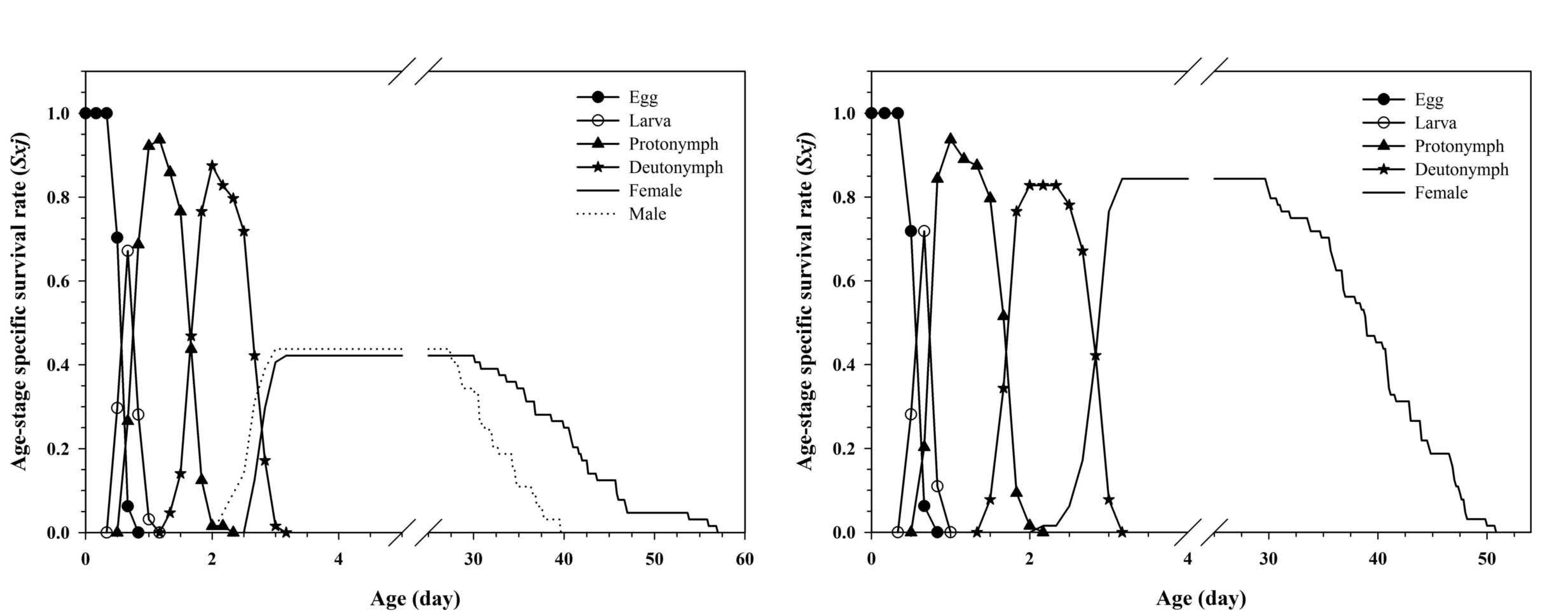
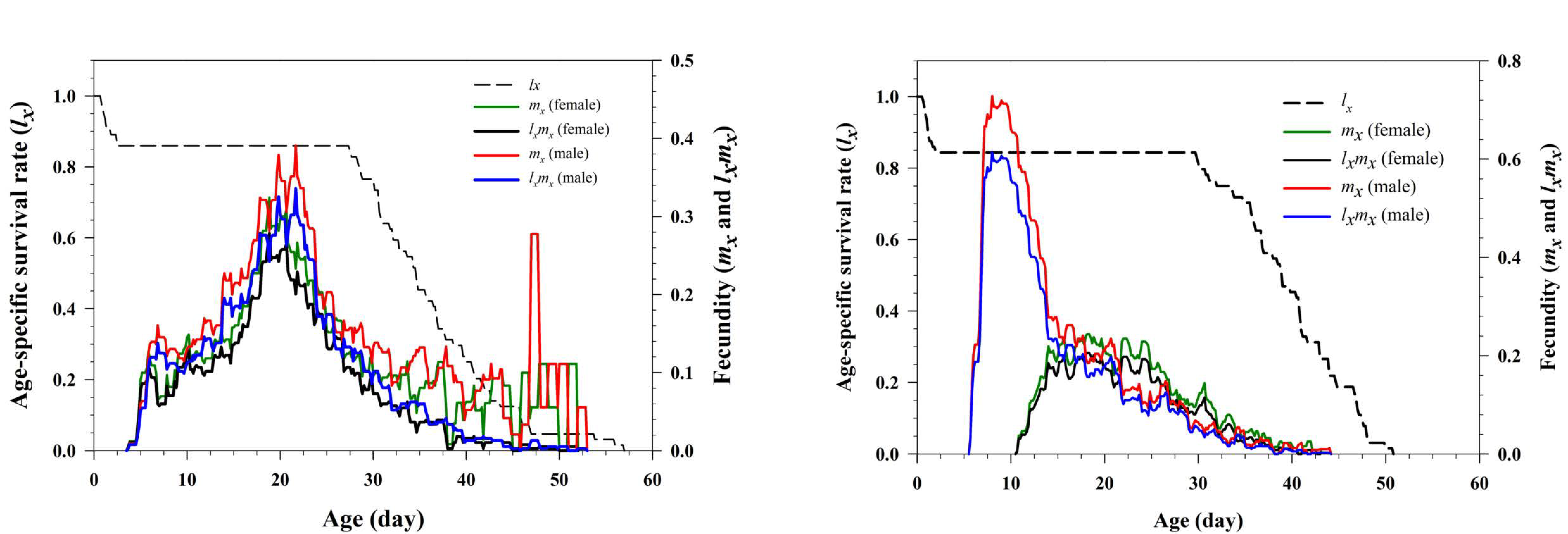
The age-specific survival rate (lx ), fecundity rate (mx ) and age-specific maternity (lxmx ) were calculated and are displayed in Figure 2. The age-specific survival rate demonstrates a simplified overview of the survival history when all stages are pooled (Figure 2). Females started to produce male and female offspring at age three days in bisexual reproduction, with the daily fecundity of producing male offspring being slightly higher than that of producing female offspring in the majority of cases (Figure 2). In oedipal reproduction, virgin females started to produce male-only offspring from age five days, while female offspring were not produced until age 10 d; which was five days after mating with their parthenogenetically produced male offspring (Figure 2).

Mean adult female longevity were 38.63 ± 0.68 and 37.48 ± 0.75 days in bisexual and oedipal reproduction, respectively. There was no significant difference between the longevity of females in either reproduction system (P > 0.05). Mean adult male longevity was 30.25 ± 1.32 days in bisexual reproduction.
Mean daily fecundity were 4.04 and 2.56 offspring in bisexual and oedipal reproduction systems, respectively. In bisexual reproduction, females produced a mean total of 128.51 ± 1.4 offspring (57.96 female and 71.07 male offspring) during their lifetime, while oedipal females produced a mean total of 68.23 ± 8.9 offspring (22.89 female and 44.35 male offspring) (Table 3). There was significant difference between the total fecundity of females in the two reproductive systems (Table 3). The daily mean fecundity of females decreased after insemination in the oedipal group, while male eggs were still being produced in large numbers (Figure 2).
Population parameters for both reproductive types of M. muscaedomesticae are presented in Table 4. In bisexual reproduction the life table parameters were calculated as: the intrinsic rate of increase (rm ) = 0.2938 females/female/day, the finite rate of increase (λ) = 1.3415 population multiplication/day, the net reproduction rate (R0 ) = 54.216 offspring/individual, the gross reproductive rate (GRR) = 77.7 offspring/individual and the mean generation time (T) = 13.5885 days (Table 4). In the oedipal cohort, by including all eggs in the analysis, values of rm (0.3420 d-1) and λ (1.4073 d-1) were significantly higher than those obtained in the bisexual cohort (P < 0.05), while the mean generation time had a smaller value than that in bisexual reproduction (P < 0.05). The GRR and R0 values did not differ significantly between the two reproductive cohorts (P > 0.05) (Table 4). By including only female eggs in the calculation of population parameters (not shown in Table 4), values of rm , λ and R0 were 0.1510 ± 0.0083 d-1, 1.1629 ± 0.0098 d-1 and 19.3107 ± 2.7509 offspring, respectively. These parameters were significantly lower than corresponding ones in the bisexual group (P < 0.05). The mean generation time (19.60 ± 0.2844 d) was significantly longer than in the bisexually reproducing group, as well as in the oedipal one by including all eggs in the analysis (P < 0.05). However, because the oedipal cohort changed its reproductive strategy to bisexual after oedipal mating, it would be invalid to calculate population parameters for this cohort. This will be explained later in the discussion.


The age-stage life expectancy curves (exj ) of M. muscaedomesticae for bisexual and oedipal cohorts are plotted in Figure 3. The life expectancy of an egg was 32.1 d in the bisexual cohort, compared to 34.2 d in the oedipal cohort. After reaching adulthood, a female’s life expectancy increased to 38.8 d in the bisexual cohort and 38.2 d in the oedipal cohort.


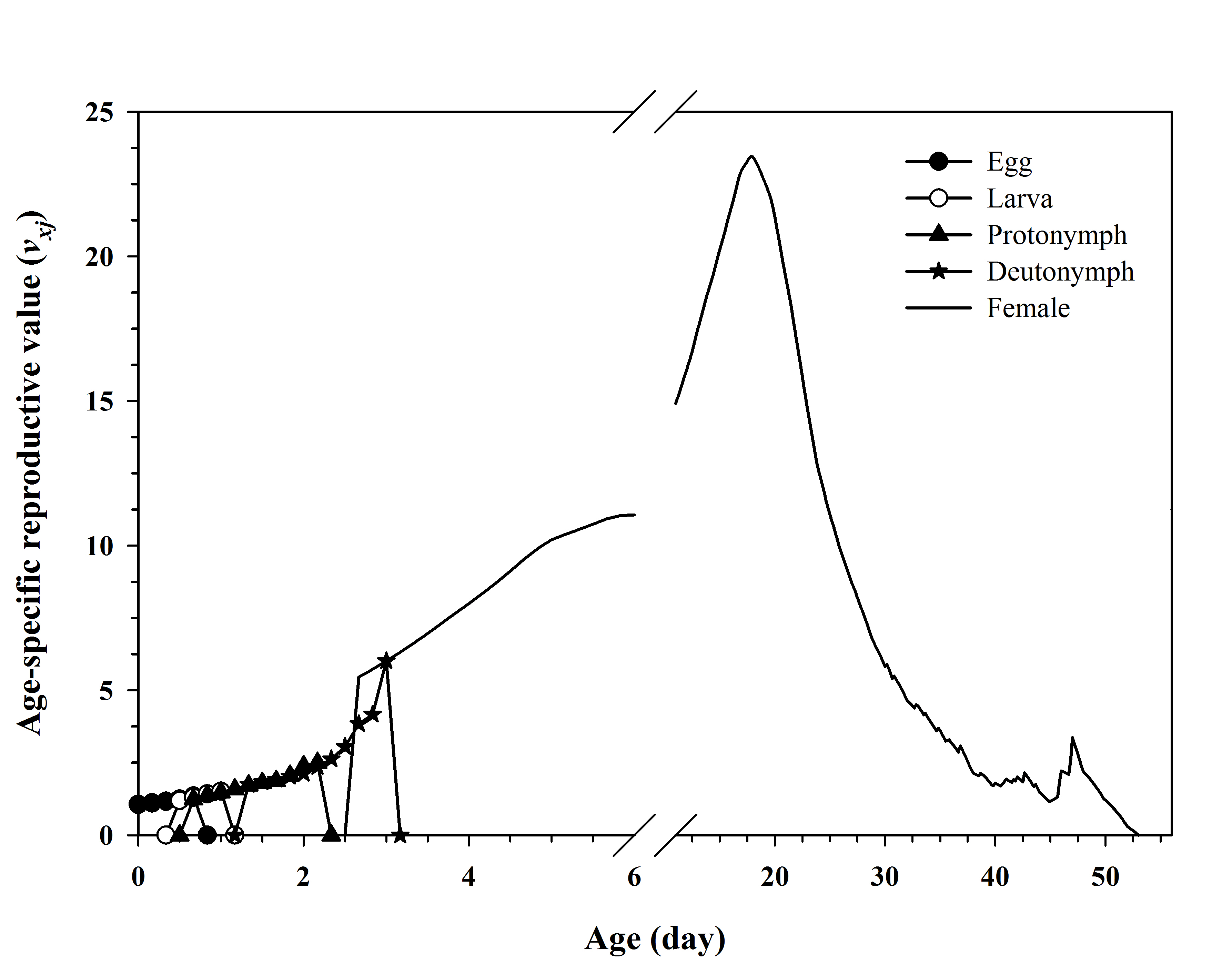
The peak reproductive value (vxj ) of the bisexual cohort occurred at age 17 d (Figure 4). The vxj curve for the oedipal cohort was not plotted because computing the reproductive value for this cohort is not applicable.

A summary of life history parameters of M. muscaedomesticae based on the present study and published literature is presented in Table 5. Accordingly, the mean egg incubation period determined in our study is similar to the findings of Abo-Taka et al. (2014), but longer than values reported by Wade and Rodriguez (1961). In our study, the highest percentage of developmental time among the juvenile stages was spent in the deutonymph stage, whereas the larval stage was the shortest period. Similarly, Wade and Rodriguez (1961) reported the deutonymph as the longest period, but the egg was the shortest stage in a bisexual population. Wade and Rodriguez (1961) and Filipponi et al., (1971) found no significant differences between the mean total preadult duration in males and females of bisexual colonies of the mite. In our study, the mean total preadult duration was slightly longer in females (68.16 h) compared to 62.88 h for males. The longest mean total preadult duration value was reported by Abo-Taka et al. (2014). The differences may be explained by disparities in experimental conditions or differences between strains of M. muscaedomesticae.


No mortality was observed in the egg stage of either studied cohort. However, M. muscaedomesticae tend to secrete their eggs in the substrate and it is possible that an occasional unnoticed egg could die in an obscure area of the experimental container and be overlooked. This possibility would likely result in the egg mortality being, at best, speculative, - a point noted by Cicolani (1979) as well.
In our study, the calculated sex ratio (females / total offspring) in the bisexual cohort (42%) was near to the findings of Wade and Rodriguez (1961), Filipponi et al. (1971) and Cicolani (1979). However, the sex ratio (females / total adults) was reported to be 75.8% in a population of M. muscaedomesticae studied by Filipponi et al. (1971) (Table 5).
The adult preoviposition period was not significantly different between mated and virgin females in the study by Wade and Rodriguez (1961), which was similar to the value observed in the bisexual cohort of our study. Based on our findings, the oviposition period of females in the bisexual cohort lasted, on average, 16% longer than females of the oedipal cohort.
According to our results, females in the bisexual group had a mean fecundity nearly two times greater than data reported by Wade and Rodriguez (1961). On the other hand, mean egg production observed by Filipponi et al. (1971) and Cicolani (1979) was higher than our figure. Mean adult female longevity was the longest in our study, and the shortest in Cicolani (1979) (Table 5).
The intrinsic rate of increase (rm ) for M. muscaedomesticae feeding on Mu. domestica eggs in this study (0.293) was lower than the only other reported value for this species (0.905) (Cicolani, 1979). Differences in several experimental variables such as prey species, strain of predatory mite, temperature, as well as calculation method may explain the lower rm value found for M. muscaedomesticae reared on Mu. domestica in our study compared to Cicolani’s results. Cicolani (1979) studied the biology and life table parameters of M. muscaedomesticae fed on a combination of Mu. domestica eggs and live rhabditid nematodes at 30°C. In addition, he stressed that due to atypical features of macrochelid biology, e.g., facultative larviparity and arrhenotoky, non-standard analyses were necessary to estimate the rm rather than the usual methods used in demographic analysis. By taking these points, particularly the last one, into consideration we believe that Cicolani overestimated the true value of rm in M. muscaedomesticae.
The rm value is a key demographic parameter for forecasting the potential success of a pest species. It can also be useful as a means of assessing promising biocontrol candidates on the basis of their reproductive potential (Roy et al., 2003). Acording to Sabelis (1992) and Sabelis et al. (2002), theoretically, a predator that has a potential growth rate equal to or greater than its prey should actively control the prey population. Nurita et al. (2012) studied the life table parameters of Mu. domestica reared on three different diets and reported rm values as 0.237, 0.245 and 0.268 (d-1) on solid waste, poultry manure and standard laboratory diet, respectively. The intrinsic rate of increase (rm ) of M. muscaedomesticae feeding on Mu. domestica eggs in the current study (0.293) is higher than the rm values reported for Mu. domestica by Nurita et al. (2012). The Iranian strain of M. muscaedomesticae, therefore, has the potential to be used as an agent for biological control of Mu. domestica in manure microhabitats.
Based on our findings, mated females produced an average of 128 offspring, which is about twice the average number produced by females in the oedipal cohort (68 offspring). The reduction of female fecundity in oedipal mating may be attributed to an inherent penalty experienced by the female in this type of mating. When males are absent, females are forced to wait to inbreed with a parthenogenetically produced son. This delay in sexual reproduction is a cost or devaluation to her fecundity (Adamson and Ludwig, 1993).
Many mites are haplodiploid and several groups have life histories that could potentially adapt mother-son mating (Adamson and Ludwig, 1993). In our study, we found that a single virgin female was capable of initiating a population through oedipal reproduction. This finding is consistent with those of Oliver (1962), Bergh (2001), McCulloch and Owen (2012) and Tuan et al. (2015) who reported oedipal reproduction in Histiostoma murchiei Hughes and Jackson (Acari: Histiostomatidae), Phyllocoptruta oleivora (Ashmead) (Acari: Eriophyidae), Ornithonyssus sylviarum (Canestrini and Franzago) (Acari: Macronyssidae) and Tetranychus urticae Koch (Acari: Tetranychidae), respectively.
Dispersal of M. muscaedomesticae can be accomplished by phoretic association with dipterans when the dung desiccates and deteriorates. Our laboratory tests showed that a combination of bisexual and oedipal reproduction can result in the founding of a new population by a single inseminated or virgin female M. muscaedomesticae. Because most female macrochelids are likely inseminated prior to dispersal, sib-mating among F1 progeny would be most relevant to the initiation of a new population. A dispersing female mite might also arrive in the midst of an established population on a new manure substrate. Alternatively, when populations are low early in the season or following a pesticide application there may be few or no conspecific males to mate with in her environment. Under such conditions, oedipal reproduction provides a means for female M. muscaedomesticae to establish a new population without being constrained by opportunities to mate.
Anderson stated in 1983 that, at that time, M. muscaedomesticae offered the greatest potential as an effective predator against all major dipteran species associated with confined poultry and livestock worldwide. He also claimed that the species exhibited the four fundamental traits of an effective natural enemy listed by Doutt and DeBach (1964), i.e., (1) high searching ability; (2) high degree of host (prey) specificity; (3) high rate of increase in relation to the pest population; and (4) ability of occupying all of the host (prey) colonized microhabitats (Anderson, 1983). In addition to having these desirable traits, our results have demonstrated that unmated female M. muscaedomesticae are also able to produce and later copulate with their sons (oedipal mating), allowing virgin females to eventually produce both sexes. In the context of predator ecology, this reproductive system is advantageous because it allows females to disperse to new manure substrates and begin colonizing in the absence of males and without prior mating. This interesting reproductive behavior considerably extends the dissemination potential of this predator, and, to some extent, helps to explain why it is as cosmopolitan and predominant a natural enemy as it is.
We are thankful to Prof. Hsin Chi (Ömer Halisdemir University, Turkey) for his help in data analysis. We also thank two anonymous reviewers for their helpful comments on an earlier draft of this manuscript. The study was financially supported by the Shahid Chamran University of Ahvaz.
Abo-Taka S.M., Heikal H.M., Abd El-Raheem A.M. 2014. Macrochelid mite, Macrocheles muscaedomesticae (Acarina: Macrochelidae) as a biological control agent against house fly, Musca domestica (Diptera: Muscidae) in Egypt. Int. J. Zool. Res., 10: 30-36. doi:10.3923/ijzr.2014.30.36 ![]()
Adamson M., Ludwig D. 1993. Oedipal mating as a factor in sex allocation in haplodiploids. Philos. Trans. R. Soc. Lond., B, Biol. Sci., 341: 195-202. doi:10.1098/rstb.1993.0103 ![]()
Akköprü E.P., Atlıhan R., Okut H., Chi H. 2015. Demographic assessment of plant cultivar resistance to insect pests: a case study of the dusky-veined walnut aphid (Hemiptera: Callaphididae) on five walnut cultivars. J. Econ. Entomol., 108: 378-387. doi:10.1093/jee/tov011 ![]()
Anderson J.R. 1983. Mites as biological control agents of dung-breeding pests: Practical consideration and selection for pesticide resistance. In: Hoy M.A., Cunningham G.L., Knutson L. (Eds) Biological control of pests by mites: University of California Press. p. 99-102.
Akca I., Ayvaz T., Yazıcı E., Smith C.L., Chi H. 2015. Demography and population projection of Aphis fabae (Hemiptera: Aphididae): With additional comments on life table research criteria. J. Econ. Entomol., 108: 1466-1478. doi:10.1093/jee/tov187 ![]()
Axtell R.C. 1961. New records of north American Macrochelidae (Acarina: Mesostigmata), and their predation rates on the house fly. Ann. Entomol. Soc. Am., 54: 748-748. doi:10.1093/aesa/54.5.748 ![]()
Axtell R.C. 1963. Acarina occurring in domestic animal manure. Ann. Entomol. Soc. Am., 56: 628-633. doi:10.1093/aesa/56.5.628 ![]()
Bergh G.C. 2001. Ecology and aerobiology of dispersing citrus rust mite (Acari: Eriophyidae) in central Florida. Environ. Entomol., 30(2): 318-326. doi:10.1603/0046-225X-30.2.318 ![]()
Chi H. 1988. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol., 17: 26-34. doi:10.1093/ee/17.1.26 ![]()
Chi H. 2017. TWOSEX-MSChart: a computer program for the age-stage, two-sex life table analysis [Internet]. Available from: http://140.120.197.173/Ecology/ ![]()
Chi H., Liu H. 1985. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sinica, 24: 225-240.
Cicolani B. 1979. The intrinsic rate of natural increase in dung macrochlid mites, predators of Musca domestica eggs. Boll. Zool., 46(3): 171-178. doi:10.1080/11250007909440296 ![]()
Doutt R.L., Debach P. 1964. Some biological control concepts and questions. In: Debach, P. (Ed). Biological control of insect pests and weeds. London, Chapman & Hall. p. 118-142.
Efron B., Tibshirani R.J. 1993. An introduction to the bootstrap. New York, Chapman & Hall, pp. 456. doi:10.1007/978-1-4899-4541-9 ![]()
Fathipour Y. 1994. Soil mites fauna in orchards of Tabriz and population fluctuation and abundance of important species [MSc thesis]. Tarbiat Modarres University of Tehran. pp. 213.
Filipponi A. 1964. The feasibility of mass producing macrochelid mites for field trials against house flies. Bull. World Health Organ., 31: 499-501.
Filipponi A., Mosna B., Petrelli G. 1971. The optimum temperature of Macrocheles muscaedomesticae as a population attribute. Riv. Parassitol., 32: 193-218.
Fisher R.A. 1930. The genetical theory of natural selection. Oxford, Clarendon Press. pp. 318.
Galazzi D., Nicoli G. 1996. Comparative study of strains of Phytoseiulus pesimilis Athias-Henriot (Acarina: Phytoseiidae). I. Development and adult life. Boll. Ist. Entomol. “G. Grandi” Univ. Bologna, 50: 215-231.
Gerson U., Smiley R.L., Ochoa R. 2003. Mite (Acari) for pest control. Oxford, Blackwell Science Ltd. pp. 539. doi:10.1002/9780470750995 ![]()
Goodman D. 1982. Optimal life histories, optimal notation, and the value of reproductive value. Am. Nat., 119: 803-823. doi:10.1086/283956 ![]()
Ho C.-C. 1985. Mass production of the predaceous mite, Macrocheles muscaedomesticae (Scopoli) (Acarina: Macrochelidae), and its potential use as a biological control agent of house fly, Musca domestica L. (Diptera: Muscidae) [PhD thesis]. University of Florida. pp. 196.
Huang Y.B., Chi H. 2012. Age-stage, two-sex life tables of Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae) with a discussion on the problem of applying female age-specific life tables to insect populations. Insect Sci., 19: 263-273. doi:10.1111/j.1744-7917.2011.01424.x ![]()
Kazemi S., Rajaei A. 2013. An annotated checklist of Iranian Mesostigmata (Acari), excluding the family Phytoseiidae. Persian J. Acarol., 2 (1): 63-157.
Keiding J. 1999. Review of the global status and recent development of insecticide resistance in field populations of the housefly Musca domestica (Diptera: Muscidae). Bull. Entomol. Res., 89 (Suppl. 1): S9-S67.
Khoobdel M., Davari B. 2011. Fauna and abundance of medically important flies of Muscidae and Fanniidae (Diptera) in Tehran, Iran. Asian Pac. J. Trop. Dis., 4(3): 220-223. doi:10.1016/S1995-7645(11)60073-4 ![]()
Koppert 2010. New soil-dwelling predatory mite very effective against thrips infestations [Internet]. Available from: https://www.koppert.com/news-biological-systems/new-soil-dwelling-predatory-mite-very-effective-against-thrips-infestations/ ![]()
McCulloch J.B., Owen J.P. 2012. Arrhenotoky and oedipal mating in the northern fowl mite (Ornithonyssus sylvarum) (Acari: Gamasida: Macronyssidae). Parasit. Vectors, 5: 281-285. doi:10.1186/1756-3305-5-281 ![]()
Nurita A.T., Abu Hassan A., Aiman Hanis J. 2012. Life tables and development of Musca domestica (Diptera: Muscidae) on three different diets. 24th International Congress of Entomology, 19-25 August, Daegu, South Korea.
Oliver J.H. 1962. A mite parasitic in the coccoons of earthworms. J. Parasitol., 48: 120-123. doi:10.2307/3275424 ![]()
Pereira C., de Castro M.P. 1945. Contribution to the knowledge of the type species of Macrocheles Latr. (Acarina): M. muscaedomesticae (Scopoli, 1772). Emend. Arq. Inst. Biol., 16: 153-186.
Perring T.M., Lackey L.J. 1989. Temperature and humidity effect on mortality and preadult development of two Phytoseiulus persimilis strains (Acari: Phytoseiidae). Int. J. Acarol., 15: 47-52. doi:10.1080/01647958908683821 ![]()
Roy M., Brodeur J., Cloutier C. 2003. Effect of temperature on intrinsic rates of natural increase (rm) of a coccinellid and its spider mite prey. Biocontrol, 48: 57-72. doi:10.1023/A:1021289832664 ![]()
Sabelis M.W. 1992. Predatory arthropods. In: Crawley M.J. (Ed) Natural enemies: The population biology of predators, parasites and disease: Oxford, Blackwell Scientific Publications, p. 225-264. doi:10.1002/9781444314076.ch10 ![]()
Sabelis M.W., van Baalen M., Pels B., Egas M., Janssen A. 2002. Evolution of exploitation and defense in plant-herbivore-predator interactions. In: Dieckmann U., Metz J.A.J., Sabelis M.W., Sigmund K. (Eds) The adaptive dynamics of infectious diseases: In Pursuit of virulence management: Cambridge, UK, Cambridge University Press, p. 279-321. doi:10.1017/CBO9780511525728.028 ![]()
Sharififard M., Mossadegh M.S., Vazirianzadeh B., Zarei-Mahmoudabadi A. 2011. Interactions between Entomopathogenic fungus, Metarhizium anisopliae and sublethal doses of spinosad for control of house fly, Musca domestica. J. Arthropod. Borne Dis., 5(1): 28-36.
Sharififard M., Safdari F. 2013. Evaluation of resistance or susceptibility of the house fly, Musca domestica L., of semi-industrial livestock farms to some pyrethroid insecticides in Ahvaz, southwestern Iran. Jundishapur J. Health. Sci., 5(3): 201- 206.
Tuan S.-J., Lin Y.-H., Yang C.-M., Atlihan R., Saska P., Chi H. 2016. Survival and reproductive strategies in two-spotted spider mites: demographic analysis of arrhenotokous parthenogenesis of Tetranychus urticae (Acari: Tetranychidae). J. Econ. Entomol., 109(2), 502-509. doi:10.1093/jee/tov386 ![]()
Wade C.F., Rodriguez J.G. 1961. Life history of Macrocheles muscaedomesticae (Acarina: Macrocheldiae) a predator of the house fly. Ann. Entomol. Soc. Am., 54: 776-788. doi:10.1093/aesa/54.6.776 ![]()
Senior Professor
University of São Paulo, ESALQ, Brazil
04 April 2018
The reader is pleasantly surprised with the authors’ creativity in coining the “oedipal reproduction” in the title of their paper. However, it is a pity, that their resourcefulness extended no further, since their results have since long being published, as in Manning & Halliday, 1994 – Australian Entomologist 21(3):89-94, easily found in Web of Science. As a matter of fact, even earlier: Pereira & Castro, 1947 – Arquivos do Instituto Biologico, São Paulo,18:71-89, clearly state “a parthenogenetic female, when left together with the males of its offspring, may become gametogenetic” !



2017-05-29
Date accepted:
2017-11-23
Date published:
2018-03-15
Edited by:
Roy, Lise

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Farahi, Sara ; Shishehbor, Parviz and Nemati, Alireza
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)