



An expanded description of the threatened tree kangaroo tick Ixodes (Sternalixodes) dendrolagi Wilson, 1967 (Acari: Ixodidae) from Papua New Guinea
Kwak, Mackenzie L.1
1✉ Department of Biological Sciences, National University of Singapore, 16 Science Drive, Singapore 117543, Republic of Singapore.
2018 - Volume: 58 Issue: 2 pages: 408-413
https://doi.org/10.24349/acarologia/20184249ZooBank LSID: C4679895-A573-4C79-8BFC-1C400CF6B948
Keywords
Abstract
Of the almost 900 tick species known, only a limited number are threatened with extinction (Mihalca et al., 2011). Three threatened tick species, including the tree kangaroo tick (Ixodes dendrolagi), are known to exist in Papua New Guinea. Hoogstraal (1982) noted the minimal research the Papuan tick fauna had attracted, and to date little is known about the tick species which are found there. Accurate identification is required to better understand the ecology of the Papuan ticks and to enhance conservation efforts for the limited number of species threatened with extinction. The recent collection of I. dendrolagi from its type host in its type locality has enabled the documentation of more morphological variation in this species than was previously recognised. This has facilitated the production of a more accurate description of the species.
Two adult female specimens, one engorged and the other unengorged, were taken from a Matschie's tree-kangaroo Dendrolagus matschiei on the Huon peninsula, Morobe district, Papua New Guinea (Figure 1) in 2014. Both specimens were deposited in the Victorian Agricultural Insect Collection (VAIC). These specimens were identified based on morphological features using the taxonomic description of I. dendrolagi provided by Wilson (1967). Morphological features were examined using both a Hitachi TM3030 Tabletop scanning electron microscope and a LEICA M205 C light microscope. Morphometric parameters, apart from idiosoma width and length in one unengorged female, were all within the range presented by Wilson (1967) and are therefore not reported. Measurements of the idiosoma were taken using a LEICA M205 C light microscopy and associated application suite and are presented in millimeters. Scanning electron microprograms are presented in Figure 2 taken using the same Hitachi TM303 Tabletop scanning electron microscope mentioned above.


Ixodes Latreille, 1795
Idiosoma
body — unengorged idiosoma ovoid, widest between spiracular plates, unengorged length from apices of scapulae to posterior body margin (3.22 mm); width (2.10 mm); engorged idiosoma oblong, widest anterior to spiracular plates; moderate number of short fine setae on ventral and dorsal surfaces, more numerous outside genital groove.
Scutum — distinctly longer than wide, widest slightly anterior to terminal edge of lateral carinae posterolaterally, anterolateral margin with slight concavity, posterior margin rounded; scapulae blunt, broadly rounded; lateral carinae distinct, slightly curved along length of scutum, beginning at inner margin of scapulae, extending two thirds length of scutum to anterolateral edge of posterolateral margin of scutum; corrugations absent on posterior margin of scutum; cervical grooves faint, deepest anteriorly, extremely faint posteriorly, terminating slightly posterior to widest part of scutum; scutum surface smooth, with short indistinct setae, inconspicuous evenly distributed punctations throughout, three uneven chains of large punctations present at anterior edge of scutum between scapulae at level of cervical grooves, several uneven chains of large punctations along posterior edge of scutum.



Sternal plate — elongate triangular, edges rounded, shape variable, surface with small, scattered punctations, small number of fine setae present on surface.
Spiracular plate — subcircular, slightly elevated; chain of aeropyles along edge of plate roughly parallel to sides, other aeropyles distributed randomly towards centre of plate, no aeropyles directly around perimeter of loosely prolate spheroid shaped spiracle; approximately as long as wide.
Genital aperture — approximately level with coxae IV in partially engorged and unengorged specimens, operculum wider than long, lateral edges slightly recurved anteriorly in engorged specimens, U-shaped in unengorged specimens, posterior margin broadly rounded.
Genital groove — anterior margin rounded with slight apical notch in engorged specimen, anterior margin strait in unengorged specimen, diverging gradually to terminate parallel with anal aperture.
Anal groove — approximately ovoid, groove meets posteriorly to form a point; anal valves with three pairs of setae. Gnathosoma
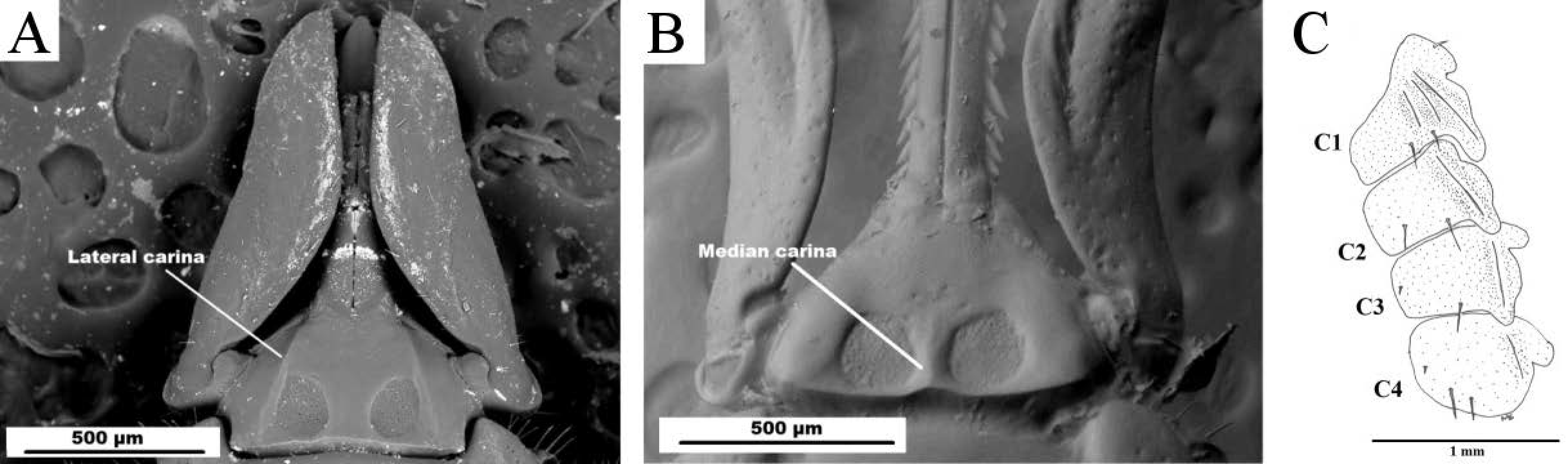
Capitulum — Basis capituli ranges from pentagonal to subpentagonal dorsally, surface crazed; median carina present, extending slightly past anterior edge of porose areas, lateral carinae absent; porose areas subcircular, large, deep, separated by approximated half their width; large punctations present anteromedian to porose areas, ranging from many to very few; posterior margin of basis capituli bilobulate; cornua absent. Basis capituli subhexagonal ventrally; auriculae blunt, incompletely retrograde extensions raised above surface of basis; carinae absent; transverse sutural line present below auriculae; posterior margin broadly rounded.
Palps — long and narrow, longer than hypostome; article I hyoid shaped ventrally, rounded; article II and III fussed, setae few and short, inconspicuous evenly distributed punctation, anterior margin rounded, outer margin concave, inner margin strongly convex, inner surface concave; article IV circular and reduced, on ventral side of palpal article III, several setae present on surface.
Hypostome — lanceolate, bluntly pointed apically; dentition 3/3 for three-fourths length, then 2/2 to base; files 1 and 2 with approximately 13 teeth, file 3 with approximately 18 teeth, forth file occasionally represented by 1 or 2 small teeth at tip; denticles of files 1 and 2 large and pointed, larger than denticles of file 3, denticles of file 3 two sizes. Legs
Coxae — large bluntly pointed external spurs present on coxae I – III, largest on coxa I, smallest on coxa III; very small external spur present or absent on coxa IV; coxa I with internal salience, internal salience absent on coxae II – IV; coxae I – III with rugulosities externally and/posteriorly, coxa IV with rugulosities present or absent, strongest on coxa I, weakest on IV (Fig. 3C); coxae I – IV with posterior margins salient.
Trochanters — trochanters I – III with ridges, best developed on II; trochanters III and IV with small, inconspicuous ventral spurs.
Tarsi and pulvili — Tarsia abruptly narrowed distally, with moderate subapical humps; pulvilli almost as long as claws.
The female I. dendrolagi can be distinguished as a member of the Sternalixodes by the presence of a sternal plate which is absent from all other described ticks outside the subgenus. Within Sternalixodes only I. dendrolagi and I. confusus exhibit a median carina on the dorsal surface of the basis capituli, between the porose areas. However, the two can easily be distinguished by the presence of lateral carinae on the dorsal surface of the basis capituli in I. confusus which are absent in I. dendrolagi. Carinae present on the basis capituli of Sternalixodes are presented in Figs. 3A-B. As I. dendrolagi does not exhibit lateral carinae on the dorsal basis capituli, this character is shown on the dorsal basis capituli of I. hirsti.
Based on examination of these new specimens, I. dendrolagi appears to exhibit wider morphological variation than previously recorded. In the original description of the species, small coxal spurs were also recorded on the forth coxae (Wilson, 1967). However, in the recently examined specimens spurs were absent from the forth coxae. The presence or absence of a coxal spur on the final coxae is also reported in I. cordifer, to which I. dendrolagi is apparently closely related (Wilson, 1967; Roberts, 1970). Rugulosities also varied somewhat in the recently collected specimens compared with the original description of I. dendrolagi (Fig. 3C). In the original description rugulosities were recorded only on coxae I–III. However, weakly developed rugulosities were also present on coxa IV in the recently collected specimens. Again this concurs with reports in other sternalixodid ticks of rugulosities, referred to as “coxal ridges” by Roberts (1970), being sometimes present or absent in coxae III and IV (Roberts, 1970). Other than these morphological variations, the specimens collected matched well with the morphological description of I. dendrolagi produced by Wilson (1967).
It has been proposed that host specific ticks may become threatened when their primary host is threatened (Mihalca et al. 2011). A tick species may even become more threatened than its host(s) if the tick’s populations are isolated due to host range fragmentation. This has been demonstrated to lead to genetic bottlenecking in Amblyomma sphenodonti, a host specific tick of the Tuatara (Sphenodon punctatus), which today occurs on only eight of the twenty eight offshore islands occupied by its host (Miller et al. 2007).
Ixodes dendrolagi has been collected only from Matschie's tree-kangaroo (Dendrolagus matschiei) and Doria's tree-kangaroo (Dendrolagus dorianus) on the Huon peninsula (Mihalca et al. 2011). Mihalca et al. (2011) designated I. dendrolagi as threatened as its known hosts D. matschiei and D. dorianus are listed as endangered and vulnerable, respectively. While the concept of parasite conservation has begun to gain traction, there remains limited information on methodologies for ensuring population stability and recovery in threatened parasite species (Dougherty et al. 2015). One potential method for safe guarding parasite species from extinction is ex situ conservation, in which populations of the parasite are maintained in captivity along with their known hosts. A captive breeding program is presently run by the Association of Zoos and Aquariums (AZA) to conserve D. matschiei (McGreevy et al. 2009). Ixodes dendrolagi could be amalgamated into this program to ensure its survival. Another potential method for preventing the extinction of I. dendrolagi is conservation translocation, in which ticks may be translocated to populations of hosts where they have become locally extinct.
I thank Professor Ian Beveridge for specimens without which this research could not have been undertaken. I also thank Dr Bruce Halliday for originally suggesting this study.
Dougherty E.R., Carlson C.J., Bueno V.M., Burgio K.R., Cizauskas C.A., Clements C.F., Seidel D.P., Harris N.C. 2015. Paradigms for parasite conservation. Conserv. Biol., 30: 724-733. doi:10.1111/cobi.12634 ![]()
Homsher P.J., Keirans J.E., Robbins R.G., Irwin-Pinkley L., Sonenshine D.E. 1988. Scanning electron microscopy of ticks for systematic studies: structure of Haller's organ in eight species of the subgenus Sternalixodes of the genus Ixodes (Acari: Ixodidae). J. Med. Entomol., 25: 348-353. doi:10.1093/jmedent/25.5.348 ![]()
Hoogstraal H. 1982. Ticks (Acari: Ixodoidea): a preliminary study. In: Gressit JL (ed) Biogeography and ecology of New Guinea. Netherlands: Springer pp. 537-544. doi:10.1007/978-94-009-8632-9_24 ![]()
Kwak M.L., Beveridge I., Koehler A.V., Malipatil M., Gasser R.B., Jabbar A. 2017. Phylogenetic analysis of the Australasian paralysis ticks and their relatives (Ixodidae: Ixodes: Sternalixodes). Parasit. Vectors., 10: 122. doi:10.1186/s13071-017-2045-4 ![]()
McGreevy T.J., Dabek L., Gomez-Chiarri M., Husband T.P. 2009. Genetic diversity in captive and wild Matschie's tree kangaroo (Dendrolagus matschiei) from Huon Peninsula, Papua New Guinea, based on mtDNA control region sequences. Zoo. Boil., 28: 183-196.
Mihalca A.D., Gherman C.M., Cozma V. 2011. Coendangered hard-ticks: threatened or threatening?. Parasit. Vectors., 4: 71. doi:10.1186/1756-3305-4-71 ![]()
Miller H.C., Conrad A.M., Barker S.C., Daugherty C.H. 2007. Distribution and phylogenetic analyses of an endangered tick, Amblyomma sphenodonti. New. Zeal. J. Zool., 34: 97-105. doi:10.1080/03014220709510068 ![]()
Roberts F.H.S. 1970. Australian Ticks. Melbourne: CSIRO Publishing. pp. 18-63.
Wilson N. 1967. Ixodes dendrolagi (Metastigmata: Ixodidae), a New Species of Tick from Tree Kangaroos (Marsupialia: Macropodidae) in New Guinea. J. Parasitol., 53: 847-851. doi:10.2307/3276786 ![]()



2017-08-07
Date accepted:
2017-11-22
Date published:
2018-02-22
Edited by:
McCoy, Karen

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Kwak, Mackenzie L.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)