



Suitability of date palm pollen as an alternative food source for the predatory mite Cydnoseius negevi (Swirski and Amitai) (Acari: Phytoseiidae) at a low relative humidity
Alatawi, Fahad J.1 ; Basahih, Jamal S.2 and Kamran, Muhammad3
1✉ Acarology Laboratory, Department of Plant Protection, College of Food & Agriculture Sciences, King Saud University, Riyadh 11451, Saudi Arabia.
2Acarology Laboratory, Department of Plant Protection, College of Food & Agriculture Sciences, King Saud University, Riyadh 11451, Saudi Arabia.
3Acarology Laboratory, Department of Plant Protection, College of Food & Agriculture Sciences, King Saud University, Riyadh 11451, Saudi Arabia.
2018 - Volume: 58 Issue: 2 pages: 357-365
https://doi.org/10.24349/acarologia/20184247Keywords
Abstract
Phytoseiid mites (Mesostigmata: Phytoseiidae) play an important role as biocontrol agents in different agro-ecosystems throughout the world (Helle and Sabelis, 1985; Gerson et al. 2003; McMurtry et al. 2013). Many phytoseiid species have been successfully introduced in crops to control phytophagous mites especially spider mites and other small insect pests (McMurtry et al. 2013). The phytoseiid mite, Cydnoseius negevi (Swirski & Amitai), is a common species in the Middle East (Abou-Awad et al. 1998; Palevsky et al. 2009; Hountondji et al. 2010). It is also widely distributed in Saudi Arabia and has been found to be abundant in date palm orchards along with the date palm mite Oligonychus afrasiaticus (McGregor) (Acari: Tetranychidae) (Negm et al. 2012a,b; Alatawi et al. 2017). However, the ability of this predator to control the date palm mite and other pests in fields has not been studied yet.
As a generalist facultative (type III) phytoseiid predator (Momen et al. 2009; McMurtry et al. 2013), C. negevi is able to successfully develop and reproduce on a wide range of food sources i.e., eriophyid and spider mites, thrips, and pollen of different plant species (Momen 1997; Momen et al. 2009; Negm et al. 2014; Hussein et al. 2016). Plant pollens contain high contents of proteins and essential amino acids and serve as high nutritional quality food for phytoseiid mites (Cook et al. 2003; Riahi et al. 2016, 2017). Therefore, plant pollens play an important role in the persistence and dynamics of many generalist predators (Hanna et al. 2005). The possibility of mass rearing a phytoseiid predator on alternative and more economical diet such as pollen could increase the interest in that predator as a biocontrol agent (Castagnoli and Simoni 1999).
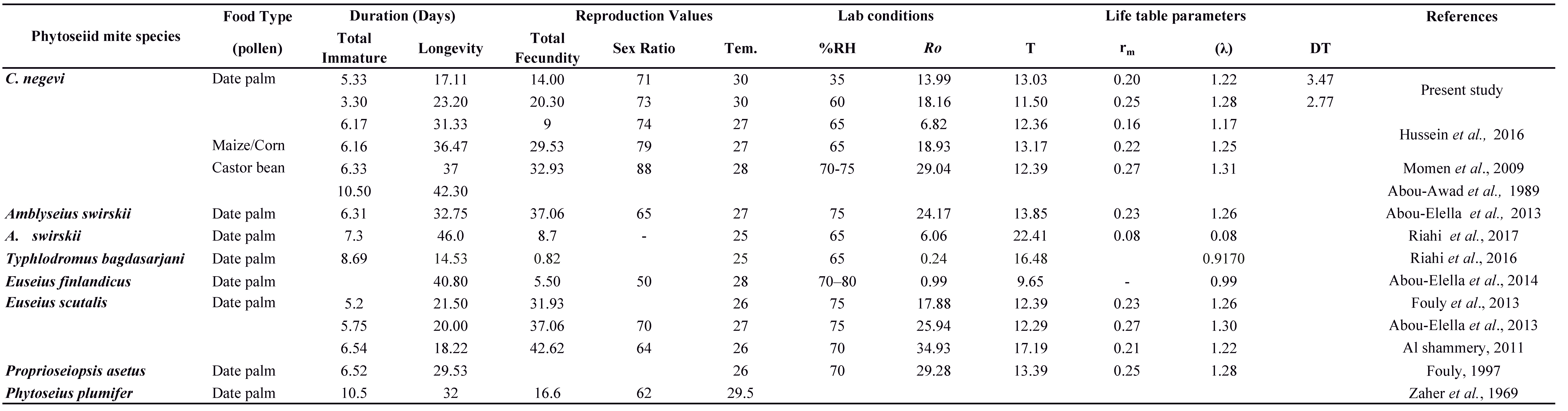
The biology and life table of C. negevi, when feeding on pollen of different plant species and on some mites and insects under different temperatures and relative humidities in the laboratory have been investigated (Tables 4 and 5). However, the biological responses of C. negevi under low humidity, the general climate of Saudi Arabia (SA), have been poorly tested.
The biology and life table parameters of several phytoseiid mite species feeding on date palm pollen have been studied so far, e.g., Amblyseius swirskii Athias-Henriot, Typhlodromus (Anthoseius) bagdasarjani Wainstein & Arutunjan (Abou-Elella et al. 2013; Riahi et al. 2016; 2017), Euseius scutalis (Athias-Henriot) (Al-Shammery 2011; Abou-Elella et al. 2013; Abou-Elella et al. 2014; Fouly et al. 2013), E. finlandicus (Oudemans) (Abou-Elella et al. 2014), and Proprioseiopsis asetus (Chant) (Fouly 1997) (Table 4). Moreover, several studies have been conducted on C. negevi feeding on different preys, e.g., tetranychid and eriophyid mites, and small insects (Table 5) (Momen 1997; Abou-Awad et al. 1989; Negm et al. 2014; Metwally et al. 2015) (Table 5). All these biological studies were conducted at high relative humidity, i.e., not less than 65 % RH.
The aim of this study was to investigate the suitability of date palm pollen as an alternate food source for C. negevi at relatively low humidity level (35 % R.H.). This study could be used in predicting C. negevi colonization ability when date palm pollen provided as alternative food source at the time of releasing the predator in date palm orchards where relative humidity is low.
Cydnoseius negevi was collected from the bermuda grass, Cynodon dactylon L. (Poaceae), under the date palm trees, Phoenix dactylifera L. at Alammariah, Riyadh, SA, 24°48.354′N, 46°29.937′E, during September 2014. The C. negevi colony was then maintained in small arenas as described by Marafeli et al. (2011), placed in an incubator in the laboratory at 27 ± 2 °C and 65 ± 10 % RH and was fed on mostly date palm pollen and rarely on the two prey species, Tetranychus urticae Koch and O. afrasiaticus. The populations of O. afrasiaticus, collected from highly infested date palm trees and T. urticae collected from egg plants at King Saud University (KSU), were maintained on different grasses and common bean, Phaseolus vulgaris L. (Fabaceae) plants respectively in a growth chamber at the Department of Plant Protection, College of Food and Agriculture Sciences, (KSU), Riyadh. Fresh date palm pollen was directly collected from male date palm trees, at KSU from March to April 2014. The open flowers of the male date palms were dried in the laboratory at 25°C and 30 % RH, shaken on a white paper, and the pollen was collected in small glass vials and stored in a refrigerator until their use (Al-Shammery 2011).
The experiment was conducted during August–September 2015 in a climate controlled chamber (BINDER®, Germany) at 30 ± 2 °C, two relative humidity levels, 35 % ± 5 and 60 % ± 5, RH, 14 L:10 D in small experimental units following Marafeli et al. (2011), with a few modifications. The experimental units consisted of a black plastic lid (3 cm diameter) placed on a petri dish (9 cm diameter) without a cover. Wet cotton wool wrapped around the lid and distilled water was provided continuously in the Petri dish to prevent the mites from escaping. Wisps of cotton wool were placed on the arena to provide oviposition sites for the predator. Seventy-four replicates for each RH level were made. Two C. negevi eggs were transferred from the stock colony to each experimental cell using a fine brush. After hatching, each larva was kept in the experimental cell separately and each fed with equal amount of date palm pollen. Older pollen was replaced with fresh date palm pollen under a stereomicroscope (SZX10, Olympus, Tokyo, Japan) using a fine brush each 3 - 4 days. A male from the stock colony was introduced to each experimental unit for 12 hours when the female deutonymph stage appeared. After mating, males were removed from the cells. The mated females were individually confined in experimental units and date palm pollen was supplied until their death. The biological parameters, i.e., eggs incubation period, duration of larval, protonymphal, and deutonymphal stages, and lifespans of female (including pre-oviposition, oviposition, post-oviposition, and fecundity) and male predators were observed and recorded at 12-h intervals until their death. The sex ratio of the progeny was also recorded.
Data were analyzed using a one way ANOVA to compare the developmental period of different immature stages, adult longevity and life span of male and female and female fecundity of C. negevi according to the two RH tested. Means were compared using Fishers LSD test at P < 0.05. All analyses were run using the SAS computer program version 9.2 (SAS Institute®, 2008).
The life history data of the adult females were used to develop the life table parameters. The life table parameters (i.e., net reproductive rate (Ro), intrinsic rate of increase (rm), finite rate of increase (λ), mean generation time (T), and doubling time (DT)) were calculated according to Birch (1948) using a software program developed by Abou-Setta et al. (1986).
Cydnoseius negevi successfully completed its development when fed on date palm pollen at both 35 % and 60 % RH levels and 30 °C. The immature developmental time, total life span, and longevity of male and female and different reproductive parameters of the female are shown in Tables 1 and 2.
The total duration of the immature stage development of both C. negevi females and males were significantly lower at 60 % RH (3.30 and 4.45 days, respectively) than at 35 % RH (5.33 and 6.01 days, respectively) (F3.70 = 16.72; P < 0.0001). At both 35 % and 60 % RH, the duration of the developmental stage of C. negevi immature males was longer than that of females (Table 1). The incubation period (F3.70 = 24.11; P < 0.0001), duration of the larval (F3.70 = 11.12; P < 0.0001) and protonymphal stages (F3.70 = 10.32; P < 0.0001) were significantly higher at 35 % RH than at 60 % RH. However, the developmental duration of the deutonymphs of both sexes was not influenced significantly at both RH levels (Table 1).

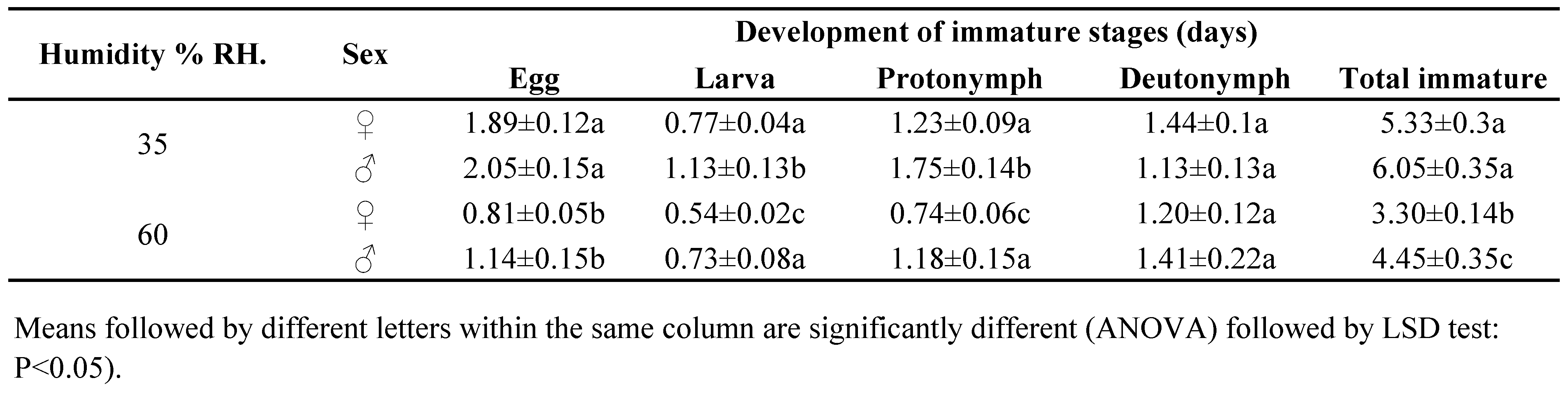
Decreasing humidity levels from 60 % to 35 % had a significant effect on the mean total fecundity and longevity of C. negevi females (Table 2). Cydnoseius negevi females deposited fewer eggs (14) at 35 % RH than at 60 % RH (20.3) (F1.57 = 7.11; P < 0.01), although the oviposition period and total life span of the females did not significantly differ (F1.57 = 3.56; P < 0.06) at the two RH levels (Table 2). The sex ratio of C. negevi progeny was female-biased and not affected by RH (71 % and 73 % of females, for 35 % and 60 % RH, respectively) (Table 2).


C. negevi females successfully reproduced when fed on date palm pollen at both 35 % and 60 % RH levels and 30 °C; life table parameters are presented in Table 3. However, when humidity decreased from 60 % to 35 % RH, “net reproductive rate (R0),” “intrinsic rate of increase (rm)” and “finite rate of increase (λ)” also decreased (Table 3). The duration of mean generation time (T) and doubling time (DT) were longer at 35 % than at 60 % RH (Table 3). At 60 % and 35 % RH, the population of this species could multiply 18 and 14 times (R0 = 18.16 and 13.99 females/female) with the generation time ranging from 11.5 to 13.05 days, respectively. Furthermore, the intrinsic rate of increase (rm) decreased from 0.25 to 0.20 individuals/female/day and the finite rate of increase (λ) decreased from 1.28 to 1.22 at 60 % and 35 % RH, respectively.


The purpose of this study was to evaluate the biology and life table parameters of C. negevi, fed on date palm pollen at a low humidity level (35 % RH). Cydnoseius negevi reproduced successfully at both 35 % and 60 % RH levels. Previous studies showed that C. negevi reproduced and developed successfully when fed on pollens of different plant species such as date palm Phoenix dactylifera L, maize, Zea mays L. (Hussein et al. 2016), and castor bean, Ricinus communis L. (Abou-Awad et al. 1989; Momen et al. 2009) at the humidity level 65 % RH (Table 4). Negm et al. (2014) also showed that C. negevi successfully preyed on the date palm mite, O. afrasiaticus, and completed its development under low relative humidity 35 % RH. The ability of C. negevi to develop and reproduce at a relatively low humidity (35 %) as well as feed on different small pests and variety of plant pollen (Table 4 and 5), could thus explain its survival in arid areas.

In the present study, eggs incubation period and the total immature developmental duration of C. negevi increased as relative humidity decreased from 60 % to 35 %. This probably explains the differences in the total duration of the immature stage development at the different humidity levels tested (Table 1). Similar trends were recorded when Typhlodromus aripo De Leon was reared under low humidity conditions feeding on cassava green mite Mononychellus progresivus Doreste, (Mutisya et al. 2014).
However, at both 35 and 60 % RH, the developmental time of C. negevi immature females feeding on date palm pollen was shorter in the present study than that of C. negevi fed on the date palm pollen (Hussein et al. 2016), pollens of castor bean and maize (Abou Awad et al. 1989; Momen et al. 2009). Likewise, when fed on date palm pollen, the immature stages of the phytoseiid mite, Euseius scutalis Athias-Henriot, completed its development in a significantly shorter time than when fed on citrus and castor bean pollens (Al-Shammery 2011). These results could indicate that date palm pollen may accelerate the development of immature stages. Results in this study showed that the developmental period of C. negevi was a little longer for immature males than for females at both 35 and 60 % RH. Similarly, the immature developmental time of the phytoseiid mites, A. swirskii and Neoseiulus californicus (McGregor) was longer for the males than for the females when fed on pollen of Betula pendula Roth and castor beans (Goleva and Zebitz 2013; Marafeli et al. 2014).
In addition, in this study, total mean fecundity and longevity of C. negevi females significantly declined when humidity levels decreased from 60 to 35 %. Similar trends were reported by Mutisya et al. (2014) while studying biology of T. aripo feeding on cassava green mite, M. progresivus Doreste. However, the total mean fecundity and adult longevity of C. negevi fed on date palm pollen, even at 60 % RH, was recorded greatly lower in the present study as compared when it fed on castor bean pollen, Ricinus communis L at 70‒75 % RH (Abou Awad et al. 1989; Momen et al. 2009) and on maize pollen, Zea mays L. at 65 % RH (Hussein et al. 2016) at almost the same temperature. Furthermore, among the different plant pollen tested, the highest fecundity and longevity were reported when C. negevi females fed on castor bean pollen (Abou Awad et al. 1989; Momen et al. 2009) (Table 4). Among all prey and diets tested (table 4 and 5), the highest total fecundity and longevity of C. negevi were recorded for females reared on the two spider mite species, T. urticae (Momen 1997).
Often, fecundity, oviposition period, and longevity of phytoseiid mites were found to be higher and longer at high humidity than at low (Kumari and Sadana 1991; Schausberger 1998; Mutisya et al. 2014) as found in this study. However, total mean fecundity values were higher in three previous studies conducted at 70-75 % RH when E. scutalis fed on date palm pollen than recorded in the present study even at high RH (65%), but the longevity found was the same (Al-Shammery 2011; Abou-Elella et al. 2013; Fouly et al. 2013) (Table 4). Interestingly, Hussein et al. (2016) found that total mean fecundity of C. negevi females fed on date palm pollen at 65 % RH was very low (9 eggs/ female for all its life) although female longevity was longer than what was observed in the present study (Table 4).
In the present study, the sex ratio (female %) was not significantly affected by humidity. Similarly, females of E. alstoniae showed insignificant difference in the sex ratio at a constant temperature (30 °C) with different relative humidity levels (50, 70 and 90 %) (Kumari and Sadana 1991). Under relatively high humidity (65–80 % RH) and different diets consumed by C. negevi, the female proportion ranged from 55 to 88 % (Table 4 and 5). In the present study, C. negevi female proportion was recorded almost the same (71 %, 73 %) at both 35 % and 60 % RH. Among the different plant pollens tested previously, the highest C. negevi female proportion was recorded when individuals were fed on castor bean pollen (88 %) (Momen et al. 2009). This suggests that diet can be one of the main factors affecting the sex ratio as pollens from different plant species have different nutritional values (Todd and Bretherick 1942; Stanley and Linskens 1974; Schmidt and Buchmann 1985; Atrouse et al. 2004; Nicolson 2011). However, Abou-Setta et al. (1997) found that the proportion of females (56–58 %) within the species Proprioseiopsis rotundus (Muma) was not affected when fed with different kinds of food.

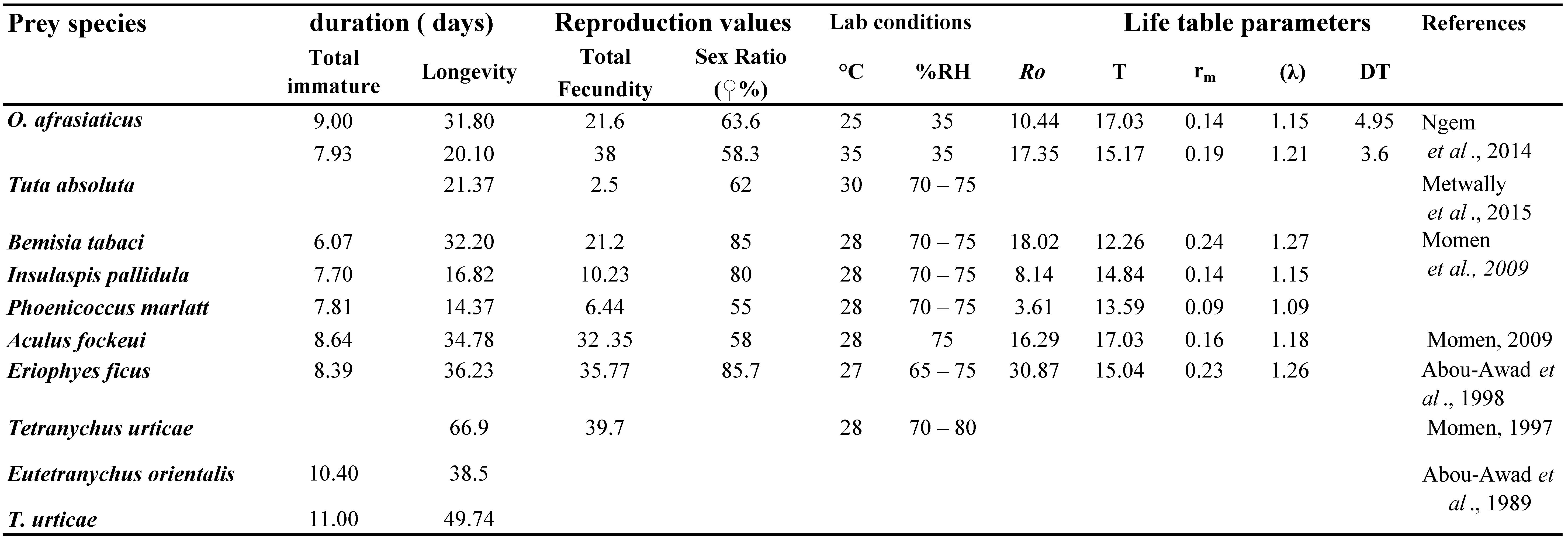
Overall, life table parameters of C. negevi were influenced negatively when humidity levels decreased from 60 to 35 %. In previous studies, life table parameters of C. negevi were only studied at high RH (>65 %) except in Negm et al. (2014) at 35 % RH (Table 4). The net reproductive rate (R0) and mean generation time (T) of C. negevi, when fed on date palm pollen were comparatively lower (13.91 and 13.03) at 35 % RH and 30 °C in this study than when fed on O. afrasiaticus (17.37 and 15.17) at same RH but 35 °C respectively (Negm et al. 2014). On the opposite, at 35 % RH, there was no significant difference in the doubling time (DT), intrinsic rate of increase (rm) and finite rate of increase (λ) of C. negevi feeding on either date palm pollen in the present study or O. afrasiaticus (Negm et al. 2014) (Table 5). Among the different plant pollens tested, including date palm pollen in the present study, the highest net reproductive rate (R0) of C. negevi was recorded when fed on castor bean pollen (Momen et al. 2009). The mean generation time (T) of C. negevi was almost the same when fed either on date palm pollen in the present study or on pollens of castor bean, maize and date palm in previous studies even at different RH levels (Momen et al. 2009; Hussein et al. 2016). However, AL-Shammery (2011) reported that the date palm pollen shortened the mean generation time of E. scutalis as compare when it fed on the pollen of other plant species i.e. citrus, alfalfa, and castor bean and simultaneously resulted in the highest values of net reproductive rates, intrinsic and finite rates of increase. Recently, the life table parameters for T. bagdasarjani and A. swirskii fed on date palm pollen were determined in two independent studies (Riahi et al. 2016; 2017) (Table 4 and 5). Both phytoseiid species exhibited longer mean generation times (T) compared to C. negevi in the present study on the same food. Also the net reproductive rate (R0) of both species when fed on date palm pollen were lower than for C. negevi in the present study. However, both species exhibited higher values for net reproductive rate (R0), intrinsic rate of increase (r), and finite rate of increase (λ) when fed on almond pollen (Riahi et al. 2016, 2017).
The date palm mite O. afrasiaticus is usually found in association with C. negevi in fields on the date palm trees and grasses (Alatawi et al. 2017). The population of date palm mite greatly decline in off date fruit period especially in winter (Palevsky et al. 2003). Therefore, the provision of date palm pollen during winter and off fruit period on date palm trees and underground grasses may enhance the population of the predators especially C. negevi. In the augmentative releases of natural enemies, the provision of pollen on the plant surface may enhance or sustain the population growth of predatory mites, which will ultimately result in enhanced biocontrol conditions (Nomikou et al. 2002).
Cydnoseius negevi successfully developed and reproduced when fed date palm pollen at a relatively low humidity (35 % RH). Thus, we would recommend it as a biocontrol agent of the date palm mite under arid environmental conditions. Moreover, date palm pollen could be used as a suitable diet for the augmentation and conservation of C. negevi in date palm orchards (grass and date palm leaves) as well as for its survival as a biocontrol agent on date fruits before early infestation of date palm mites. When N. californicus (a generalist predator) is released on perennial crops prior to spider mite infestation and in the presence of artificial food (pollen), with exceptions, it could persist and maintain the pest population at a level lower than the economic threshold level (Schausberger and Walzer 2001).
Further field-based studies are required for a better understanding of the potential use of C. negevi with date palm pollen in the field where systems are likely to be more complex than laboratory conditions. No study on the release of this predator has been applied in the field yet.
The authors would like to extend their sincere appreciation to the Deanship of Scientific Research at the King Saud University for funding this research (Research Group no. RG 1437-043). Also, we would like to thank Prof. Mohamed Abou-Setta (Plant Protection Research Institute, Egypt) for his guidance in calculating life table parameters using BASIC 48 program.
Abou-Awad B.A., El-Sherif A.A., Hassan M.F., Abou-Elela M.M. 1998. Laboratory studies on development, longevity, fecundity and predation of Cydnoseius negevi (Swirski & Amitai) (Acari: Phytoseiidae) with two mite species as prey. J. Plant Dis. Protect., 105: 429-433. Available from http://www.jstor.org/stable/43215262 ![]()
Abou-Awad B.A., Nasr A.K., Gomaa E.A., Abou-Elela M.M. 1989. Life history of the predatory mite, Cydnodromella negevi and the effect of nutrition on its biology (Acari: Phytoseiidae). Insect Sci. Appl., 10: 617-623. doi:10.1017/S1742758400021743 ![]()
Abou-Elella G.M., Hassan M.F., Nawar M.S., Zidan I.M. 2014. Survival, development and reproduction of Euseius finlandicus (Oudemans) (Acari: Phytoseiidae) fed on various kinds of food substances. Arch. Phytopathol. Plant Protect., 47(7): 857-868. doi:10.1080/03235408.2013.823715 ![]()
Abou-Elella G.M., Saber S.A., El-Sawi S.A. 2013. Biological aspects and life tables of the predacious mites, Typhlodromips swirskii (Athias-Henriot) and Euseius scutalis (Athias-Henriot) feeding on two scale insect species and plant pollen. Arch. Phytopathol. Plant Protect., 46(14): 1717-1725. doi:10.1080/03235408.2013.774715 ![]()
Abou-Setta M.M., Fouly A.H., Childers C.C. 1997. Biology of Proprioseiopsis rotendus (Acari: Phytoseiidae) reared on Tetranychus urticae (Acari: Tetranychidae) or pollen. Fla. Entomol., 80: 27-34. doi:10.2307/3495973 ![]()
Abou-Setta M.M., Sorrell R.W., Childers C.C. 1986. Life 48: a BASIC computer program to calculate life table parameters for an insect or mite species.
Fla. Entomol., 69: 690-697. doi:10.2307/3495215 ![]()
Alatawi F.J., Basahih J., Kamran M. 2017. The superfamily Phytoseioidea (Acari: Mesostigmata) from Saudi Arabia: a new species, new records and a key to the reported species. Acarol., 57(2): 275-294. doi:10.1051/acarologia/20164154 ![]()
Al-Shammery K.A. 2011. Plant pollen as an alternative food source for rearing Euseius scutalis (Acari: Phytoseiidae) in Hail, Saudi Arabia. J. Entomol., 8: 365-374. doi:10.3923/je.2011.365.374 ![]()
Atrouse O.M., Oran S.A., Al-Abbadi S.Y. 2004. Chemical analysis and identification of pollen grains from different Jordanian honey. Int. J. Food Sci. Technol., 39: 413-417. doi:10.1111/j.1365-2621.2004.00798.x ![]()
Birch L.C. 1948. The intrinsic rate of natural increase of an insect population. J. Animal Ecol., 17: 15-26. doi:10.2307/1605 ![]()
Cook S.M., Awmack C.S., Murray D.A., Williams I.H., 2003. Are honey bees' foraging preferences affected by pollen amino acid composition. Ecological Entomology, 28(5), pp.622-627.
Castagnoli M., Simoni S. 1999. Effect of long-term feeding history on functional and numerical response of Neoseiulus californicus (Acari: Phytoseiidae). Exp. Appl. Acarol., 23(3), pp.217-234.
Fouly A.H. 1997. Effects of prey mites and pollen on the biology and life tables of Proprioseiopsis asetus (Chant)(Acari, Phytoseiidae). J. Appl. Entomol., 121(1-5): 435-439. doi:10.1111/j.1439-0418.1997.tb01431.x ![]()
Fouly A.H., Nassar O.A., Osman M.A. 2013. Biology and Life tables of Esieus scutalis (A.-H.) reared on different kinds of food. J. Entomol., 10: 199-206. doi:10.3923/je.2013.199.206 ![]()
Gerson U.L., Smiley R., Ochoa R. 2003. Mites (Acari) for pest control.Blackwell Science Ltd.: Malden, MA, USA. pp. 539. doi:10.1002/9780470750995 ![]()
Goleva I., Zebitz C.P. 2013. Suitability of different pollen as alternative food for the predatory mite Amblyseius swirskii (Acari, Phytoseiidae). Exp. Appl. Acarol., 61(3): 259-283. doi:10.1007/s10493-013-9700-z ![]()
Hanna R., Onzo A., Lingeman R., Yaninek J.S., Sabelis M.W. 2005. Seasonal cycles and persistence in an acarine predator-prey system on cassava in Africa. Pop. Ecol., 47(2): 107-117. doi:10.1007/s10144-005-0215-2 ![]()
Helle W., Sabelis M.W. 1985. Spider mites: their biology, natural enemies and control. Elsevier: Amsterdam, Netherlands. pp. 790.
Hountondji F.C., Moraes G.J.D., Al-Zawamri H. 2010. Mites (Acari) on coconut, date palm and associated plants in Oman. Syst. Appl. Acarol., 15(3): 228-234. doi:10.11158/saa.15.3.7 ![]()
Hussein H.E., El-Ela M.A., Reda A.S. 2016. Development, survival and reproduction of Typhlodromus negevi (Swirski and Amitai) (Acari: Phytoseiidae) on various kinds of food. Egypt J. Biol. Pest Control., 26(1): 43. Available from http://search.proquest.com/openview/25ed8b7cd8db7d677df10bf0cd944506/1?pq-origsite=gscholar&cbl=886351 ![]()
Kumari M., Sadana G.L. 1991. Influence of temperature and relative humidity on the development of Amblyseius alstoniae (Acari: Phytoseiidae). Exp. Appl. Acarol., 11(2): 199-203. doi:10.1007/BF01246091 ![]()
Marafeli P.D.P., Reis P.R., Silveira E.C., Toledo M.A., Souza-Pimentel G.C. 2011. Neoseiulus californicus (McGregor, 1954) preying in different life stages of Tetranychus urticae Koch, 1836 (Acari: Phytoseiidae, Tetranychidae). Acarologia, 51(4): 499-506. doi:10.1051/acarologia/20112031 ![]()
Marafeli P.P., Reis P.R., Silveira E.C., Souza-Pimentel G.C., Toledo M.A. 2014. Life history of Neoseiulus californicus (McGregor, 1954) (Acari: Phytoseiidae) fed with castor bean (Ricinus communis L.) pollen in laboratory conditions. Braz. J. Biol., 74(3): 691-697. doi:10.1590/bjb.2014.0079 ![]()
McMurtry J.A., De Moraes G.J., Sourassou N.F. 2013. Revision of the lifestyles of phytoseiid mites (Acari: Phytoseiidae) and implications for biological control strategies. Syst. Appl. Acarol., 18(4): 297-320. doi:10.11158/saa.18.4.1 ![]()
Metwally A.M., Momen F.M., Nasr A.K., Abdallah A.A., Ebadah I.M., Saleh K.M. 2015. Prey suitability of Tuta absoluta larvae (Lepidoptera: Gelechiidae) for three predatory phytoseiid mites (Acari: Phytoseiidae) under laboratory conditions. Acta Phytopathol. Entomol. Hung., 50(1): 105-113. doi:10.1556/038.50.2015.1.10 ![]()
Momen F.M. 1997. Copulation, egg production and sex ratio in Cydnodromella negevi and Typhlodromus athiasae (Acari, Phytoseiidae). Anz Schadl., 70(2): 34-36. https://link.springer.com/article/10.1007/BF01991955 ![]()
Momen F., Abdel-Khalek A., El-Sawi S. 2009. Life tables of the predatory mite Typhlodromus negevi feeding on prey insect species and pollen diet (Acari: Phytoseiidae). Acta Phytopathol. Entomol. Hung., 44(2): 353-361. doi:10.1556/APhyt.44.2009.2.12 ![]()
Mutisya D.L., El-banhawy E.M., Kariuki C.W., Khamala C.P.M. 2014. Typhlodromalus aripo De Leon (Acari: Phytoseiidae) development and reproduction on major cassava pests at different temperatures and humidities: an indication of enhanced mite resilience. Acarologia, 54: 395-407. doi:10.1051/acarologia/20142145 ![]()
Negm M.W., Alatawi F.J., Aldryhim Y.N. 2012a. A new species of Neoseiulus Hughes, with records of seven species of predatory mites associated with date palm in Saudi Arabia (Acari: Phytoseiidae). Zootaxa., 3356: 57-64. Available from https://www.researchgate.net/publication/227645301 ![]()
Negm M.W., Alatawi F.J., Aldryhim Y.N. 2012b. Incidence of predatory phytoseiid mites in Saudi Arabia: new records and a key to the Saudi Arabian species (Acari: Mesostigmata: Gamasina). Syst. Appl. Acarol., 17(3): 261-268. doi:10.11158/saa.17.3.6 ![]()
Negm M.W., Alatawi F.J., Aldryhim Y.N. 2014. Biology, predation, and life table of Cydnoseius negevi and Neoseiulus barkeri (Acari: phytoseiidae) on the old world date mite, Oligonychus afrasiaticus (Acari: Tetranychidae). J. Insect Sci., 14(1): 177. doi:10.1093/jisesa/ieu039 ![]()
Nicolson S.W. 2011. Bee food: the chemistry and nutritional value of nectar, pollen and mixtures of the two. Afr. Zool., 46(2): 197-204. doi:10.1080/15627020.2011.11407495 ![]()
Nomikou M., Janssen A., Schraag R., Sabelis M.W. 2002. Phytoseiid predators suppress populations of Bemisia tabaci on cucumber plants with alternative food. Exp. Appl. Acarol., 27(1-2): 57. doi:10.1023/A:1021559421344 ![]()
Palevsky E., Gal S., Ueckermann E.A. 2009. Phytoseiidae from date palms in Israel with descriptions of two new taxa and a key to the species found on date palms worldwide (Acari: Mesostigmata). J. Nat. Hist., 43(27-28): 1715-1747. doi:10.1080/00222930902969484 ![]()
Riahi E., Fathipour Y., Talebi A.A., Mehrabadi M. 2017. Linking life table and consumption rate of Amblyseius swirskii (Acari: Phytoseiidae) in presence and absence of different pollens. Ann. Entomol. Soc. Am., 110: 244-253. doi:10.1093/aesa/saw091 ![]()
Riahi E., Fathipour Y., Talebi A.A., Mehrabadi M. 2016. Pollen quality and predator viability: life table of Typhlodromus bagdasarjani on seven different plant pollens and two-spotted spider mite. Syst. Appl. Acarol., 21(10): 1399-1412. doi:10.11158/saa.21.10.10 ![]()
SAS Institute. 2008. The SAS System for Windows, Version 9.2 Users Guide. SAS Institute Inc., Cary, North Carolina.
Schausberger P., Walzer A. 2001. Combined versus single species release of predaceous mites: predator–predator interactions and pest suppression. Biol. Control., 20(3): 269-278. doi:10.1006/bcon.2000.0908 ![]()
Schausberger P. 1998. The influence of relative humidity on egg hatch in Euseius filandicus, Typhlodromus pyri and Kampimodromus aberrans (Acari, Phytoseiidae). J. Appl. Entomol., 122(1-5): 497-500. doi:10.1111/j.1439-0418.1998.tb01534.x ![]()
Schmidt J.O., Buchmann S.L. 1985. Pollen digestion and nitrogen utilization by Apis mellifera L. (Hymenoptera: Apidae). Comp. Biochem. Physiol. A., 82(3): 499-503. doi:10.1016/0300-9629(85)90423-2 ![]()
Stanley R.G., Linskens H.F. 1974. Pollen: Biology, chemistry and management. Springer-Verlag, Berlin; New York, USA. 307pp. doi:10.1007/978-3-642-65905-8 ![]()
Todd F.E., Bretherick O. 1942. The composition of pollens. J. Econ. Entomol., 35(3): 312-317. doi:10.1093/jee/35.3.312 ![]()
Zaher M.A., Wafa A.K., Shehata K.K. 1969. Life history of the predatory mite Phytoseius plumifer and the effect of nutrition on its biology (Acarina: Phytoseiidae). Entomol. Exp. Appl., 12(4): 383-388. doi:10.1111/j.1570-7458.1969.tb02534.x ![]()



2017-06-17
Date accepted:
2017-11-11
Date published:
2018-02-22
Edited by:
Tixier, Marie-Stéphane

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Alatawi, Fahad J.; Basahih, Jamal S. and Kamran, Muhammad
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)