



A new species-group with new species of the genus Lasioseius (Acari: Mesostigmata: Blattisociidae) associated with neotropical hispine beetles in furled leaves of Heliconia
Moraza, Maria L.1 and Lindquist, Evert E.2
1✉ Universidad de Navarra, Facultad de Ciencias, Departamento de Biología Ambiental, Campus Universitario, 31080, Pamplona España.
2Canadian National Collection of Insects, Arachnids, and Nematodes, Science & Technology Branch, Agriculture and Agri-Food Canada, Ottawa, ON, K1A 0C6, Canada.
2018 - Volume: 58 Issue: 1 pages: 62-98
https://doi.org/10.24349/acarologia/20184227ZooBank LSID: 54AA8F0F-32F0-4AD1-8D49-FB1088DD2E37
Keywords
Abstract
A remarkable diversity of mesostigmatic mites have been found to coexist with Neotropical hispine beetles in furled leaves of Heliconia and related plants. Some represent a newly described family (Lindquist & Moraza 2014), and others, new genera of Melicharidae (Moraza & Lindquist 2015, 2016). Here, we describe a new species-group of the blattisociid genus Lasioseius based on adults and immatures of four newly described and one undescribed species associated with hispine beetles of the genera Cephaloleia and Chelobasis in Costa Rica. Beetles of these Neotropical genera often coexist and undergo their life histories together in the same furled leaves of Heliconia and other Zingiberales plants (Staines 2009, Schmitt & Frank 2013, Staines & Garcia-Robledo 2014). The associations of these mites and beetles are currently being studied by Garcia-Robledo and colleagues (e.g., Garcia-Robledo et al. 2013, 2015), and a need for descriptions, names, and identification keys for these mites is relevant to those studies. Although included in the key, one species is left undescribed, due to the available exemplars at hand being too few in number and in poor morphological condition. However, the description of the species-group accounts for its attributes. Lasioseius is the most diverse and species-rich genus in the family Blattisociidae. Keys to 156 species were presented by Christian & Karg (2006), and some 195 species were recorded in the catalogue by Moraes et al. (2016). Yet, the real biodiversity of this genus remains unaccounted for. For example, based on the findings of the Arthopods of La Selva (ALAS) study in the 1990’s, approximately 50 species of Lasioseius were collected, with slide preparations made of adults, from a wide variety of habitats in just one small area of lowland tropical rainforest in Costa Rica (Lindquist 2001). Most to nearly all of those species, other than those belonging to the species-group being described here, remain undescribed. Moreover, many of those species present remarkably distinctive morphological structures of the gnathosoma and legs, indicating specialized ways of life.
Living mites were discernible by hand lens on the hispine beetles when first removed from the unfurled leaves of the host plants at the La Selva Biological Station, Heredia Province, Costa Rica in 1994. Methods of collecting and observing living mites at the ALAS laboratory were the same as presented in our previous paper concerning these beetles and their associated mites (Moraza & Lindquist 2015). Mites along with their beetle hosts were preserved in 80% ethanol, with the mites subsequently mounted into Hoyer’s medium on microslides and sealed with Glpt insulating varnish. Taxonomic concepts of the superfamilies of mesostigmatic mites, especially the Phytoseioidea and its component families, and the genera of Blattisociidae, follow those of Lindquist et al. (2009) and Lindquist and Moraza (2010, 2012). Morphological observations, measurements, and illustrations were made using compound microscopes equipped with differential interference contrast and phase contrast optical systems, drawing tubes and stage-calibrated eyepiece micrometers. Setal notation for the idiosoma follows Lindquist & Evans (1965) as modified slightly by Lindquist (1994). Measurements of structures are given in micrometers (μm), indicating the ranges among specimens measured. Dorsal shield lengths are midline from anterior margin anterior to bases of vertex setae j1 to caudal margin posterior to bases of clunal setae J5. Lengths of ventral idiosomatic shields are midline, from the anterior margin to posterior edge of each structure, including the hyaline anterior extension of the epigynal shield and the posterior cribrum of the ventrianal shield. Sternal and sternitigenital shield widths are narrowest, between bases of legs II for the former, and sequentially between bases of legs II, III, IV for the latter. Notation for leg and palpal setation follows Evans (1963, 1964, 1969). Leg lengths are from base of the coxa to apex of the tarsus, excluding the pretarsus. As the forms of the leg segments and their chaetotaxy and form of setae are so similar among the species being described, complete illustrations are provided for the legs of only one species, but figures are provided for the tarsi of legs II and IV for each species. Distinction of pore-like structures on the idiosomal integument as either poroids (lyrifissures) or glandular openings (solenostomes), as distinguished morphologically by Athias-Henriot (1969, 1971b, 1975) and physiologically by Krantz & Redmond (1987), is presented stylistically in the illustrations; gland pores are shown in circular form, while poroids are shown in elliptical form. Notation for pore-like structures of the peritrematal region follows that of Johnston & Moraza (1991).
Identifications of the hispine beetles associated with the mites were made by Project ALAS parataxonomists Danilo Brenes and Ronald Vargas, or by Derek Johnson (see acknowledgments). Some of the identities of the beetles (and background plant substrate) presented for each mite species are not in accord with current molecular studies, which indicate that there is a complex of multiple cryptic species with similar morphology in the La Selva area (Carlos Garcia-Robledo, personal communications, June 2014). These issues are treated, as necessary, in a paragraph on host identifications following description of each mite species.
The holotypes and some of the paratypes of the new species are deposited in the Instituto Nacional de Biodiversidad (INBio) of Costa Rica, Santo Domingo de Heredia; additional paratypes of each new species are deposited in the Canadian National Collection of Insects and Arachnids (CNCI), Science & Technology Branch, Agriculture & Agri-Food Canada, Ottawa, and the Museum of Zoology, University of Navarra (MZUNAV), Pamplona, Spain.
Both subgenera and species-groups have been proposed by various authors for the genus Lasioseius. Five subgenera were proposed by Christian & Karg (2006), four of which were found to be based on unreliable homoplastic attributes, as reviewed by Moraza & Lindquist (2011), and synonymized under the nominate subgenus. Their subgenus Lasioseius (Endopodalius), for which Lindquist (1964) and Lindquist & Evans (1965) earlier had recognized the scutalis species-group, is well-based apomorphically, and constitutes one of the two subgenera of Lasioseius recognized by Moraza & Lindquist (2011) and included in their key to genera and subgenera of the subfamily Blattisociinae. Within the subgenus Lasioseius (Lasioseius) sensu Moraza & Lindquist, three species-groups have been proposed by various authors, based to some extent on distinctive apomorphic attributes. The phytoseioides- group was initially recognized by Lindquist (1964) and further characterized by de Moraes et al. (2015) to include about twenty named species, many of which remain inadequately described. The porulosus- group was first recognized by Walter & Lindquist (1997) to include seven species. The floridensis- group was proposed by Mineiro et al. (2009) and includes four described species. In contrast, Christian & Karg (2006) recognized five "species-complexes", all of which are based on shades of differences of adult female structures, rather than on apomorphic attributes, e.g., whether the metapodal plates are enlarged to some extent or not; whether the ventrianal shield has four versus five or six setae other than the circumanal setae; whether leg I is shorter or longer than the idiosoma; whether some of the setae on tarsus IV are long enough to be considered macrosetae; and whether the forms of dorsal shield setae are distinctively tricarinate or not. As a result, their species-complexes suffer from the same problems as their subgenera, by including species which could be grouped more meaningfully on the basis of other, more clearly apomorphic attributes, and are not recognized here. For example, Lasioseius americanus Chant, 1963, placed in their inguinalis-complex of their subgenus Lasioseius, shares other, more distinctively apomorphic attributes that are useful in redefining their subgenus Endopodalius, including the unusual convex form of the posterior margin of sternal shield and the peculiarly penicillate shape of setae Z4 or Z5. Based on those and other attributes, L. americanus was designated as a member of the scutalis species-group by Lindquist (1964) and Lindquist & Evans (1965), and is a member of the subgenus L. (Endopodalius) sensu Moraza & Lindquist (2011).
Here, we propose a new species-group, the cassidini-group, within the subgenus Lasioseius (Lasioseius), that is apomorphically and ecologically distinctive in its association with hispine beetles in rolled leaves of Heliconia and other Zingiberales plants. We include it in a tentative key to species-groups that we currently recognize within the subgenus L. (Lasioseius), preceding a key to species of the cassidini-group itself. Although based at least in part on apomorphic attributes, such species-groups may represent different hierarchic levels among the many species in the subgenus, such that one group may eventually be shown to be a subset of another. Unquestionably, further studies will recognize other species-groups or even additional subgenera, for which names such as Aceoseius Sellnick (1941) and Hyattella Krantz (1962) may already be available.
Diagnosis, Lasioseius cassidini species-group. Dorsal shield setae collectively simple, imperceptibly or vestigially tricarinate (homoplasy); setae j2 aligned transversely with para-vertical poroids idj1 nearly at level of setae j1 and z1 (apomorphy); humeral setae r3 erect, relative to adjacent setae r2 and r4 (plesiomorphy). Palptarsus with a hyaline anterolateral flap extending from base of apotele to base of cluster of apical setae (apomorphy). Peritremes short of reaching vertex area, extending at most to level slightly beyond setae s1 (homoplasy). Male subcapitulum dimorphic in having a pair of short fimbriated processes medial to and separate from corniculi (homoplasy).
With the diagnostic attributes of genus and subgenus Lasioseius (Lasioseius) sensu Moraza & Lindquist (2011), but restricted and augmented as follows:
Idiosomatic dorsum — Dorsal shield setae collectively simple (imperceptibly or vestigially tricarinate), more elongated posterior setae sometimes slightly, sparsely barbed; setae j2 aligned transversely with paravertical poroids idj1, nearly at level of setae j1 and z1; dorsal shield holotrichous, setae z3 present, humeral setae r3 erect, r5 on shield, r6 on or off shield of females and R1 on or off shield of males; idiosoma with setae j1-j6, z1-z6, s1-s6, r2-r6, J1-J5, Z1-Z5, S1-S5, R1-R6 always present (seta in R7 position designated as ZV5). Dorsal shield with complement of 23 pairs of discernible pore-like structures (10 podonotal, 13 opisthonotal), of which seven pairs (four podonotal, three opisthonotal) superficially appear secretory (gland pores) and 16 pairs (six podonotal, 10 opisthonotal) non-secretory (poroids).
Idiosomatic venter — Adult female. Tritosternum with laciniae pilose, their fused section and columnar base without elaborations. Presternal region lacking an unpaired platelet at base of tritosternum. Sternal shield entire, fully integrated with endopodal extensions between legs I-II and II-III, with posterior margin straight or slightly concave; apex of endopodal extension between legs I-II with gland pore gbv; shield with three pairs of setae (st1-st3) and two pairs of poroids (iv1-iv2); st1 and iv1 sometimes inserted in a less sclerotized region of shield. Setae st4 and poroids iv3 on tiny metasternal platelets or on soft cuticle. Endopodal strips by legs III-IV slender, distant from sternal shield. Epigynal shield with broadly to narrowly rounded hyaline anterior margin barely or not overlapping posterior edge of sternal shield. Postgenital furrow with transverse strip of four narrow platelets. Ventrianal shield with circumanal setae and usually four pairs of ventral setae, or rarely three pairs if JV1 on soft cuticle; anal opening relatively small; postanal seta clearly longer than paranal setae. Soft opisthogastric integument with two pairs of small, sometimes partly coalesced metapodal platelets, four or five pairs of setae (ZV1, ZV3, ZV5, JV5, sometimes JV1, ZV4), and three or four pairs of poroids, flanked by posteriormost two pairs of R-setae; setae ZV4 sometimes absent, JV4 always absent; glandular pores gv3 on or off shield posterolateral to anal opening. Minor duct of sperm access system without a distant secondary enlargement.
Adult male. Base of tritosternum biarticulate; other aspects of tritosternum and presternal region as in female, and with separate sternitigenital, ventrianal, peritrematal shields. Ventrianal shield capturing metapodal sigillae anterolaterally, with circumanal setae and usually seven pairs of ventral setae (rarely six pairs if JV5 on soft cuticle), and two or three pairs of poroids; glandular pores gv3 on margin of shield at level slightly behind posterior margin of anal opening; aspects of circumanal setae, anal opening and cribrum as in female; poroids ivo3 on or off shield laterad setae ZV3. Soft opisthogastric integument with one (ZV4) or two pairs of setae (ZV5 present or absent, JV4 absent), and one or two pairs of poroids; setae ZV5 often aligned with posterior flanking R-setae, so as to resemble R7. Postanal furrow present.
Lateropeltidal region — Peritrematal shields fused with dorsal shield at level of setae s1 in females and level of r2-r3 in males; pore-like elements ips, gp3, ip3 on peritrematal shield posterior to stigma; peritremes well developed but reaching at most to level slightly beyond setae s1, well short of paravertical setae z1. Exopodal strip continuous and separate from peritrematal plate alongside coxae II-III, its extension between coxae I and II nearly touching apex of endopodal extension from sternal or sternitigenital shield.
Gnathosoma — Gnathotectum anterior margin denticulate, either truncate or with three branches of similar length. Chelicerae with movable digit usually tridentate in female, unidentate in male, with weakly fimbriated basal sheath; fixed digit with offset small subapical tooth, a row of 7-28 fine teeth of similar form along entire masticatory surface, and minute pilus dentilis; dorsal seta simple; lateral hyaline rim along paraxial face of fixed digit smooth, lacking serration at level of base of movable digit. Male spermatodactyl digitiform, projecting freely, generally anteroventrally from movable chela, not elaborated in shape. Corniculi normal in hornlike form, entire, well sclerotized and separated from base to apex, lacking a paraxial process basally. Internal malae somewhat longer than corniculi, divided subapically into a shorter, lateral, fimbriated, bluntly pointed extension and a longer, medial, smooth, slender, sharply pointed one. Adult males with pair of short fimbriated processes medially beside corniculi, separate from internal malae. Subcapitulum with seven moderately wide, transverse rows of deutosternal denticles, the anterior six rows connected by lateral edges, seventh row free. Subcapitular setae simple in form, with hp1 sometimes attenuated. Palpi normal in form and setation as described for Gamasina by Evans (1964); internal setae al of femur and al-1, al-2 of genu strongly spatulate distally. Apical anterolateral margin of palptrochanter with or without one to several spinelike structures. Palptarsus with hyaline anterolateral flap extending from base of apotele to base of cluster of apical setae (Fig. 4G).
Legs — Leg I not elongated (about as long as dorsal shield); tarsus I with biarticulated pretarsus with normally developed claws. Leg IV approximately as long as dorsal shield, and with no setae differentiated as macrosetae by being more erect and longer than tibia. Tarsi II-IV without elongated whiplike setae; apical setal processes (d-1) shorter or nearly as long as length of pretarsus to base of claws; ventral setae (v-1) similar in form, or av-1 longer and thicker than pv-1; seta md slightly more slender and attenuated, and slightly the longest of tarsal setae, or subequally so with (d-2); ventral apical process smoothly triangular, tapered to a blunt tip or with a wide, untapered, serrated apical margin; pretarsi with paradactyli symmetrically developed, or with anterior one asymmetrically larger on tarsi III and IV. Complement of setae on segments of legs I to IV holotrichous for genus Lasioseius as presented by Lindquist and Evans (1965); chaetotactic formulae for genua I-II-III-IV: (2 – 3/2, 3/1 – 2) (2 – 3/1, 2/1 – 2) (2 – 2/1, 2/1 – 1) (2 – 2/1, 3/0 – 1); tibiae: (2 – 3/2, 3/1 – 2) (2 – 2/1, 2/1 – 2) (2 – 1/1, 2/1 – 1) (2 – 1/1, 3/1 – 2). Legs II-IV of males sometimes with dimorphism evident in form of particular setae.
Immature instars — Descriptive notes for the larva and protonymph are based on one or two specimens of only one species, so they cannot be assumed to hold for members of the species-group as a whole. They are presented in the description of L. serripes.
Remarks — Although based on material representing only five species, four of which are described here, this species-group is anticipated to include a multitude of species, in view of evident preferences among host beetles and the tremendous diversity of these beetles in the Neotropics and other tropical regions.
Morphological aspects — Unless overlooked among some other members of the genus Lasioseius and other genera of Blattisociidae, the hyaline flap associated with the palptarsal apotele seems independently derivative to the new species-group. A similar structure, termed a ‘hyaline scale’, is found at the base of the palptarsal apotele, and characteristic of all members of the family Veigaiidae (Lindquist et al. 2009). It does not extend as far apically along the anterior edge of the palptarsus in veigaiids. A much smaller structure, termed a ‘small hyaline lobe’ has been noted as present or absent at the base of the palptarsal apotele among species of the melicharine genera Proctolaelaps and Mucroseius (Lindquist & Hunter 1965, Lindquist & Wu 1991).
Although characteristic of males of the new species-group, the presence of a pair of short fimbriated processes medially beside the corniculi, separate from the internal malae, has been noted and illustrated previously (Lindquist 1971) for the male of an unrelated species of Lasioseius, L. corticeus Lindquist. Therefore, this dimorphism is either homoplasious or apomorphic at a higher group level within the genus Lasioseius.
In contrast to the diversity of shapes evident among many other species of Lasioseius (e.g. Walter & Lindquist1989, 1997), the forms of male spermatodactyls and female spermathecae are notably similar among the described members of this species-group. This may argue for a relatively recent diversification of species in this group or for other behavioral aspects that prevent species introgressions among coexistent species.
Within the cassidini-group, the absence or presence and number of spinelike structures on the apical anterolateral margin of the palptrochanter is a newly-noted, specific attribute. A few SEM images indicate the presence of such structures among some other species of Lasioseius (e.g., figs. 19E, G of Walter & Lindquist 1989), but their diagnostic usefulness in other groups of the genus is problematical.
Two aspects of leg IV of the protonymph, for which we have a single specimen of just one species, merit attention as possible species-group apomorphic attributes. First, the tarsus of leg IV differs from that of legs II and III in having seta av-1 distinctively erect, longer and slightly thicker than all other tarsal setae (Fig. 12G). Except for the one undescribed species at hand, this is unlike its form on tarsus IV of the adult (no deutonymphs of this species were available for study), on which it is similar to that of tarsi II and III (Figs. 13C, 14B). As noted in the key to species, the adult female of the undescribed form at hand has seta av-1 erect, and longer and thicker than other adjacent setae, though not less adjacent ones of the tarsus (Fig. 12H). This may be an ontogenetic retention of the aforementioned protonymphal condition of tarsus IV to adulthood. Such distinctiveness of protonymphal tarsus IV has not been noted for any other species of Lasioseius or other genera of the Blattisociidae, and we are not aware of something similar to this among other families of Gamasina. Second, the rudimentary presence of seta av-1 on femur IV and the full presence of seta pl-1 on genu IV are ontogenetic accelerations of setae that are typically first expressed in the deutonymph (Evans 1963). Although ontogenetic leg chaetotactic data are sparse for other species of the genus Lasioseius (e.g., Lindquist 1971, Britto et al. 2011) and for the family Blattisociidae as a whole (e.g., Lindquist 2003, Lindquist & Moraza 2016), such acceleration of usually deutonymphal setae has not been noted for other blattisociids (EEL, personal observations).
Increased descriptive and illustrative details are given to the form of setae on tarsi II to IV of members of the cassidini-group. Based on our experience in dealing with forms that are similar to previously described species, with less detailed descriptions and illustrations, we are often left uncertain as to their identities. Moreover, as species of the cassidini-group are to be involved in further studies at a more refined level, using molecular approaches to analyze for haplotypes and dietary preferences of the mesostigmatic mites associated with the two genera of rolled-leaf hispine beetles, we hope that a more detailed morphological approach will approximate those further findings.
For a key to the genus and subgenera of Lasioseius, see the key to the world genera and subgenera of the subfamily Blattisociinae by Moraza and Lindquist (2011).
1. Dorsal shield lacking vertical (j1) and paravertical (z1) setae, leaving setae j2 as anteromedial-most pair, slightly behind level of paravertical poroids idj1; posterior region of dorsal shield with 10 or 11 pairs of setae, each of J, Z, S series with less than five pairs of setae; peritremes with anterior apices curving posteromedially above level of setae j2; female ventrianal shield with six pairs of opisthogastric setae; antiaxial face of fixed cheliceral digit with a subapical pointed process
...... phytoseioides-group sensu Lindquist, 1964 and de Moraes et al., 2015
— Dorsal shield with setae j1 and usually z1 present, inserted at transverse level of poroids idj1 anterior to setae j2; posterior region of dorsal shield with 13 to 15 pairs of setae, at least the Z-series and usually all series complete with five pairs of setae each; peritremes with anterior apices not curving posteromedially where ending at level beside or short of setae z1; female ventrianal shield with two to six pairs of opisthogastric setae; antiaxial face of fixed cheliceral digit usually lacking a subapical process
...... 2
2. Dorsal shield flanked by only two or three pairs of setae (marginals r5, R1 and ventrolateral ZV5) on soft cuticle; dorsal shield with 16 pairs of setae, post-humeral pair r4 absent; tritosternum with laciniae fused for more than half their length; genua of legs I, II, III with 12, 9 or 10, 7 or 8 setae, respectively, each lacking one or two ventral setae
...... porulosus-group sensu Walter and Lindquist, 1997
— Dorsal shield flanked by five to twelve pairs of setae (elements of marginals r5-6, R1-6, submarginals UR, and ventrolateral ZV5) on soft cuticle; dorsal shield with 18 to 23 pairs of setae, usually including post-humeral pair r4; tritosternum with laciniae free for usually more than half their length; genua of legs I, II, III usually with 13, 11, 9 setae, respectively, each usually including holotrichous set of 3, 2, 2 ventral setae
...... 3
3. Dorsal shield setae collectively simple (imperceptibly or vestigially tricarinate); setae j2 nearly aligned transversely with j1 and z1; peritremes reach at most to level slightly beyond setae s1 (but not to paravertical setae z1); palptarsus with hyaline anterolateral flap extending from base of apotele to base of cluster of apical setae
...... cassidini-group
— Dorsal shield setae variable in form, but with at least setae j1, r3, Z5 usually tricarinate or tricarinate-pectinate; setae j2 inserted clearly behind transverse level of setae j1 and z1; peritremes variable in extent, but usually reaching to insertions of paravertical setae z1, anteriorly beyond setae s1; palptarsus lacking hyaline flap near base of apotele
...... 4
4. Dorsal shield with post-humeral seta r4 short, simple, similar to r2, and not more than half as long as tricarinate seta s4 neighboring it medially; most dorsal setae tricarinate and moderately long, nearly as long as longitudinal intervals between their bases; anterior pair of sternal setae on weakly sclerotized presternal area, just before anterior margin of sternal shield; sternal shield with anteromedial patch of reticula; tarsus II with one midlateral seta, pl-2, elongated, reaching nearly to base of pretarsus
...... floridensis-group sensu Mineiro et al., 2009
— Dorsal shield with seta r4 usually similar in weakly to strongly tricarinate form and over half as long as adjacent seta s4; dorsal setae variable in form and lengths; anterior pair of sternal setae on or off well sclerotized surface of sternal shield; sternal shield variably ornamented; tarsus II with or without one or two attenuated setae
...... many other species and undescribed groups of subgenus L. (Lasioseius)
1. Ventrianal shield with 3 pairs of ventral setae, JV1 on soft cuticle (Fig. 5E); sternal shield reticulate over entire surface; setae st4 and poroids iv3 on soft cuticle (Fig. 5E); metapodal platelets connected or coalesced on each side (Fig. 5E); tarsi II-IV with ventroapical process truncate-spiculate apically (Figs 13G, 14B)
...... L. serripes n. sp.
— Ventrianal shield with 4 pairs of ventral setae, JV1 on shield (Fig. 8D); sternal shield smooth, or weakly lineate along lateral margins; setae st4 and usually poroids iv3 on metasternal platelets; metapodal platelets separate on each side; tarsi II-IV with ventroapical process bluntly pointed, smooth apically (Figs 13H, 14A, C, D)
...... 2
2. Gnathotectum with three elongated tines (Fig. 8C); fixed chela with 9-13 teeth (Fig. 8E); smaller mites, dorsal shield length less than 470 μm
...... L. fuscina n. sp.
— Gnathotectum with three short broad tines or irregularly truncate-serrate; fixed chela with 18-28 fine teeth (Fig. 10G); larger mites, dorsal shield length more than 530 μm
...... 3
3. Setae Z5, S5 thickened, blunt-tipped in distinction to other attenuated dorsal setae (Fig. 10A, B); gnathotectum with three short broad tines (Fig. 10D); poroids iv3 and setae st4 together on slender metasternal platelets (Fig. 10C)
...... L. duobtusisetis n. sp.
— Setae Z5, S5 not conspicuously thicker or blunter than other dorsal setae; gnathotectum truncate-serrate (Fig. 1D); poroids iv3 alone on soft cuticle or with setae st4 on slender metasternal platelets
...... 4
4. Sternal setae st4 on tiny metasternal plates, leaving sternal poroids iv3 on soft cuticle (Fig. 1C); sternal setae st1 on weakly sclerotized presternal region (Fig. 1C); podonotal region of dorsal shield with 23 pairs of setae, including r6; tarsus IV with distal seta av-1 ca 1.3 longer than pv-1 and 2.5 times longer than slender setae (l-1) (Fig. 14A)
...... L. cassidini n. sp.
— Sternal poroids iv3 with sternal setae st4 on slender metasternal platelets (as in Fig. 8D); sternal setae st1 on anterior margin of sternal shield; podonotal region of dorsal shield with 22 pairs of setae, r6 on soft cuticle; tarsus IV with distal seta av-1 enlarged, erect, 2.5 times longer than pv-1 and 4 times longer than short, slender setae (l-1) (Fig. 12H)
...... Lasioseius sp. (undescribed) ex Chelobasis bicolor
ZOOBANK: EB77E489-1EB6-4120-9375-44E765ABEFF3 ![]()

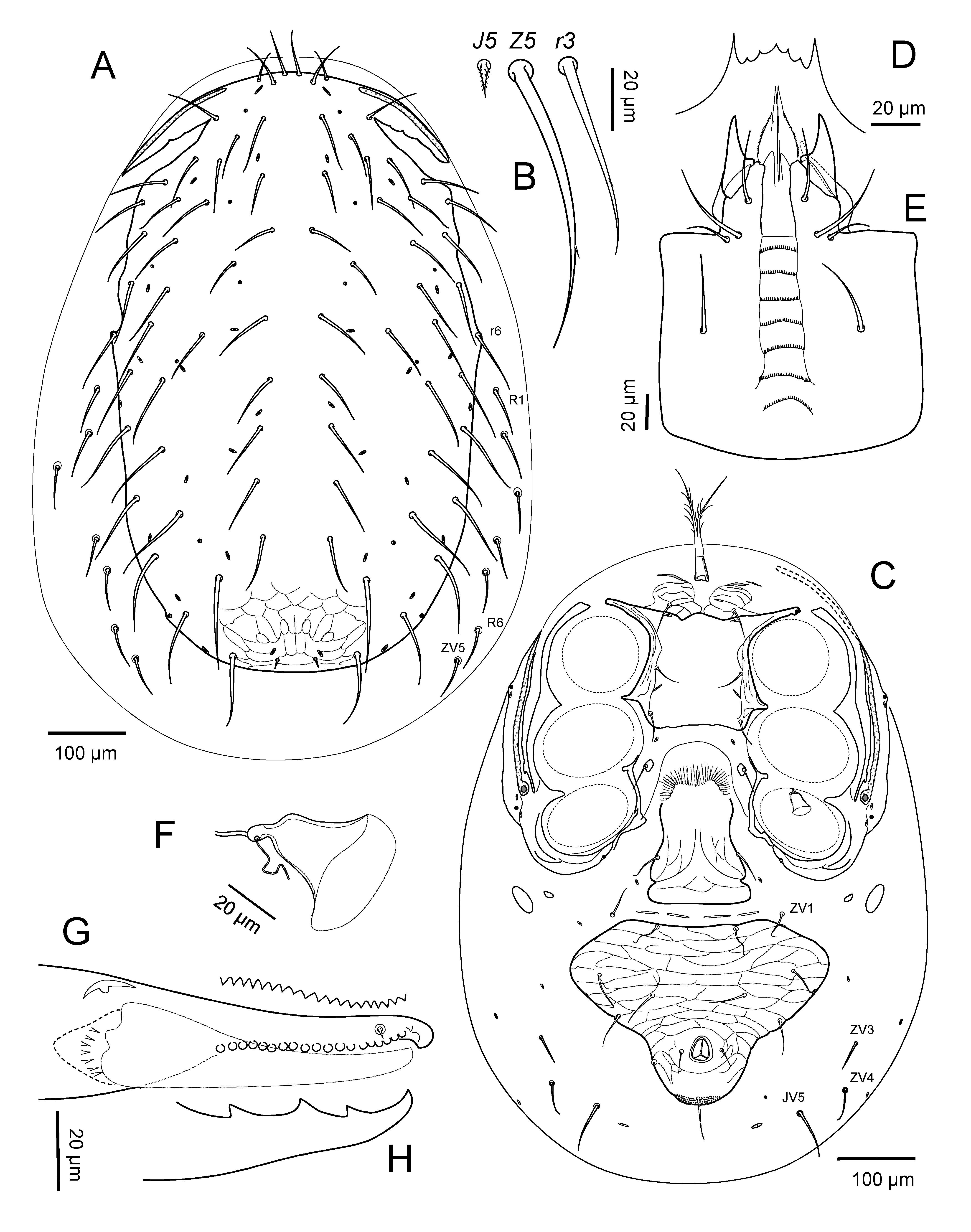

Diagnosis — Adult female: Sternal shield weakly lineate along lateral margins, smooth medially, its posterolateral corners not emarginated; poroids iv3 alone, on soft cuticle, setae st4 on tiny metasternal platelets; epigynal shield partially reticulated, its lateral margins concave, narrowed anterior to setae st5; ventrianal shield with posterolateral margins on either side of anus divergent anteriorly, and with four pairs of opisthogastric setae, JV1 on shield; metapodal plates well separated on each side. Dorsal setae r6 on edge of dorsal shield; Z5, S5 attenuated like adjacent setae. Adult male: Dorsal shield setae S5 dimorphically less attenuated. Sternitigenital shield incompletely reticulated, nearly smooth medially. Tibiae III-IV with setae (v) dimorphically spinelike, blunt; genu IV av similarly so. Adult attributes: Gnathotectum anterior margin truncate, irregularly denticulate. Fixed chela with file of ca 20 teeth in female, ca 18 in male. Sternal setae st1 inserted in weakly sclerotized presternal region. Tarsi II-IV with ventral apical process bluntly pointed, smooth apically; tarsi II-III with seta av-1 slightly longer than pv-1; pretarsi III-IV with anterior paradactylus asymmetrically elongate, twice as long as claws; tarsus IV with distal seta av-1 clearly larger, thicker than adjacent setae.
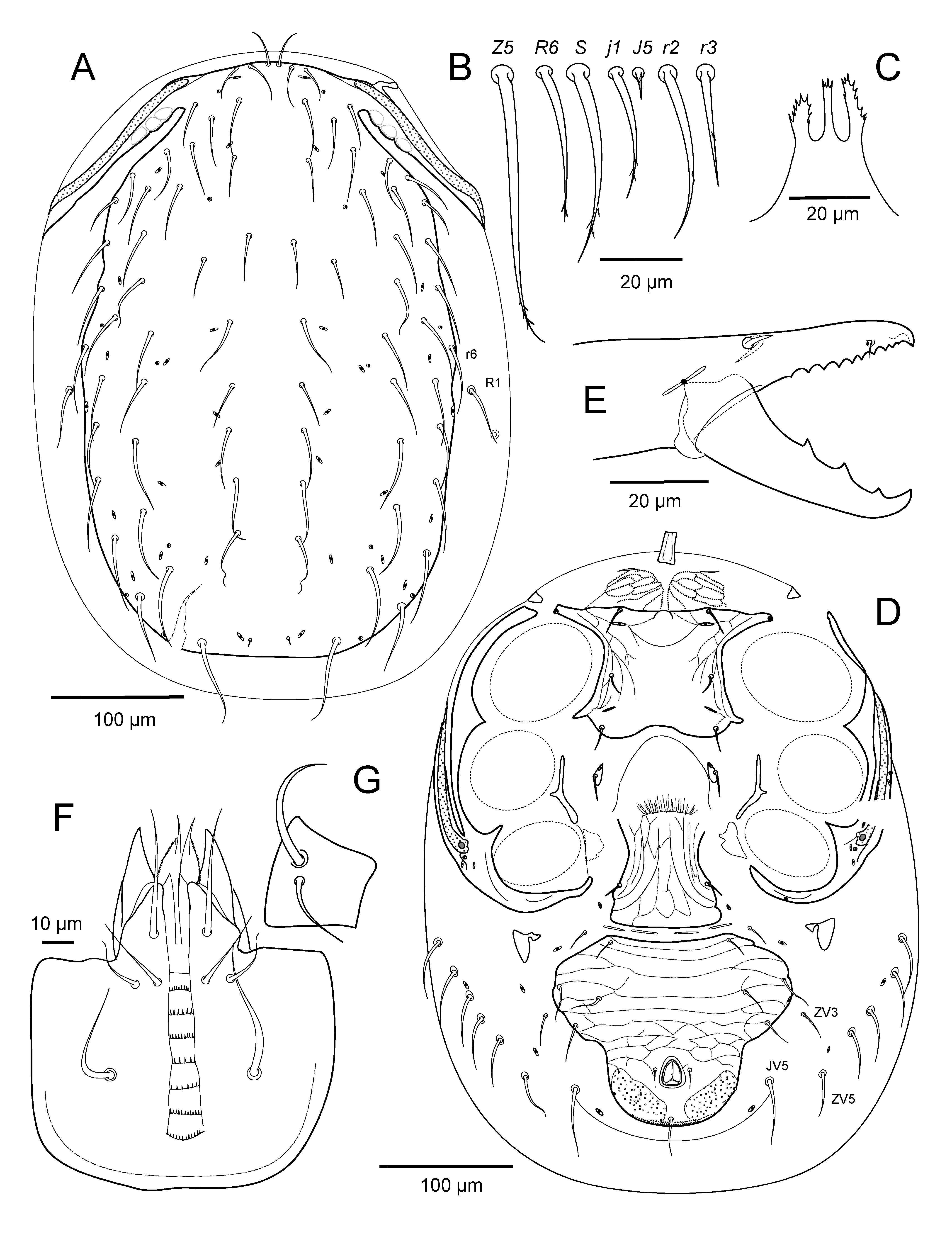
Idiosomatic dorsum — Adult female. Dorsal shield 703 – 757 long, 375 – 410 wide at level of r3, 428 – 477 wide at r6 (n=9), lightly reticulated over entire surface, without puncta behind level of setae Z4; shield with 38 pairs of setae generally similar in size and acutely pointed form, J1-J4 about as long or slightly longer than longitudinal intervals between their bases; J5 minute, barbed (8 – 12): j1, j2 (45 – 60), j3 (67 – 76), j4 (50 – 57), j5 (56 – 62), j6 (70 – 77), z1 (37 – 42), z2 (56 – 65), z3 (60 – 68), z4 (72 – 77), z5 (52 – 62), z6 (62 – 74); s1 (58 – 75), s2 (65 – 75), s3 (70 – 75), s4 (70 – 80), s5 (82 – 92), s6 (85 – 96), r2 (62 – 74), r3 erect (60 – 67), r4 (75 – 80), r5 (75 – 82), r6 (70 – 79), J1 (65 – 75), J2 (68 – 77), J3 (67 – 82), J4 (70 – 75), Z1, Z2 (92 – 103), Z3 (100 – 112), Z4 (107 – 122), Z5 (93 – 110), S1 (77 – 95), S2, S3 (88 – 103), S4 (97 – 110), S5 (82 – 105). Lateral soft cuticle with seven pairs of smooth setae, including R1-R3 (48 – 62), R4-R6 (42 – 53) and ZV5 (47 – 55) posteriorly (Figs. 1A, B, C).
Adult male. Dorsal shield 493 – 610 long, 315 – 342 wide at level of setae r3, 345 – 367 wide at r6 (n = 8); shield ornamentation, complement of pore-like structures and setation as in female; posterolateral setae attenuated as in female. Setae j1 (40 – 50), j2 (35 – 47), j3 (50 – 60), j4 (38 – 45), j5 (43 – 47), j6 (53 – 62), z1 (28 – 38), z2, z3 (43 – 55), z4 (52 – 62), z5 (40 – 45), z6 (48 – 55), s1 (47 – 57), s2-s4 (50 – 65), s5, s6 (60 – 75), r2 (50 – 60), r3 erect (38 – 55), r4, r5 (53 – 67), r6 (45 – 57), J1, J2 (43 – 55), J3 (50 – 62), J4 (40 – 47), J5 (5 – 9), Z1 (63 – 80), Z2 (65 – 92), Z3 (66 – 98), Z5 (61 – 77), S1 (54 – 72), S2 (55 – 77), S3, S4 (60 – 78), S5 (50 – 70), Z4 longest (80 – 102) of opisthonotal setae. Lateral soft cuticle with five or six pairs of R- setae (33 – 48) (Fig. 2A), R2 sometimes on small platelet (Fig. 2E).
Idiosomatic venter — Adult female. Tritosternum base sometimes biarticulate, 38 – 43 long and 20 – 22 wide at its base, with paired laciniae fused for about 22 – 25 of entire length (122 – 135). Presternal region with circumscribed pair of weakly granulated, lineated areas, flanked anteriorly by a pair of weak or distinct narrow strips on either side of tritosternal base, and with setae st1 (33 – 37) inserted on this region, anterior to sternal shield. Sternal shield 127 – 135 long from anterior margin at transverse level with poroids iv1 to slightly concave posterior margin, 125 – 136 wide, weakly lineated along lateral margins, smooth medially; sternal setae st2, st3 of similar length (30 – 37). Sternal poroids iv3 alone on soft cuticle, setae st4 (28 – 35) on tiny metasternal platelets (13 – 15 x 8 – 11). Endopodal strips narrow between coxae III and IV. Epigynal shield sparsely reticulated, its broadly rounded hyaline anterior margin not overlapping posterior edge of sternal shield, its lateral margins slightly concave, narrowed anterior to setae st5 (38 – 47), and its posterolateral corners emarginated behind st5; shield 208 – 225 long, 118 – 137 at widest level at posterior rounded corners, 80 – 87 at narrowest width between legs IV. Opisthosomatic venter with two pairs of well separated metapodal platelets, primary pair elongate-oval, 33 – 40 long, 15 – 19 wide, secondary pair small, 10 – 16 by 5 – 7. Ventrianal shield inversely subtriangular with angulose or rounded anterior corners, with posterolateral margins on either side of anus divergent anteriorly; surface entirely reticulate, and punctate on either side of postanal seta; shield wider (293 – 337) than long (212 – 232), width at anal level (130 – 155); poroids ivo2 on shield anterior edge behind setae ZV1; gland pore gv3 on or off shield posterior to anal opening; shield with four pairs of ventral setae: JV1 (37 – 42), JV2 (43 – 50), JV3 (44 – 55), ZV2 (35 – 48); with paranal setae (21 – 27) shorter than postanal seta (43 – 65). Soft cuticle with five pairs of simple opisthogastric setae, ZV1 (35 – 44) anterior to ventrianal shield, ZV3 (33 – 43), ZV4 (37 – 44), JV5 (55 – 67) flanking the shield, and ZV5 aligned behind R6; three pairs of poroids on soft cuticle. Peritrematal-exopodal shield with lines extending from stigma into rounded posterior margin (Fig. 1C); peritreme extending to level between setae s1 and z1 (Fig. 1A). Spermathecal apparatus with cup-like calyx 28 – 29 long to embolus, 30 – 31 including it (Fig. 1F).
Adult male. Tritosternum much as in female, but with base biarticulate, longer (20 – 32) than wide (15 – 19), and with laciniae fused for only about 10 – 15 of entire length (88 – 107). Presternal region faintly sclerotized, with a pair of distinct narrow strips on either side of tritosternal base, and with setae st1 inserted on this region on either side of genital opening. Sternitigenital shield length 225 – 267, narrowest widths 88 – 113 at level between coxae II, 77 – 95 between coxae III, 58 – 87 between coxae IV; shield slightly lineated along lateral margins and posteriorly between coxae IV, unornamented medially, with endopodal apices freely acuminate beside coxae IV; setae st1 (28 – 32) and st5 (28 – 37) longer, st2-st4 (17 – 26); poroids iv3 sometimes hardly discernible flanking faint pair of pits (Fig. 2A). Ventrianal shield well reticulated and with punctate fields in anal region as in female, its convex anterior margin sometimes excised near apices of exopodal strips behind coxae IV, and posterolateral margins slightly constricted in anal region; shield much wider (278 – 337) than long (170 – 228, including cribrum ca. 8 – 11 long, 58 – 65 wide); shield with six or usually seven pairs of opisthogastric setae, JV5 asymmetrically on or off shield (Figs. 1A, B); JV1 and ZV1 (23 – 32), JV2 (28 – 37), ZV2 (28 – 32), JV3 (28 – 42), ZV3 (23 – 30), JV5 (39 – 50); postanal seta smooth (39 – 47), about twice as long as paranal setae (15 – 22). Soft opisthogastric cuticle with two pairs of setae, ZV4 (26 – 30), and ZV5 (30 – 42) aligned behind R6, one pair of lyrifissures, ivp.
Gnathosoma — Adult female. Gnathotectum anterior margin broadly truncate, with two lateral smooth prongs and serrate medial edge (Fig. 1D). Cheliceral shaft, excluding basal section, 160 – 173 long, with slender digits; dorsal seta blunt (13 – 14); fixed digit with tiny offset subapical tooth and row of 18-22 very fine teeth (Fig. 1G), pilus dentilus minute, at level between 4th- 5th apical teeth; movable digit (71 – 75) tridentate (Fig. 1H). Subcapitulum rows of deutosternal denticles progressively slightly wider posteriorly, each with numerous (ca 30-35) fine denticles, 6th and sometimes 7th widest and with ca 40-45 and 35-40 teeth, respectively (Fig. 1E); subcapitular setae hp1 (40 – 45) slightly shorter than hp3 (48 – 52) and capitular seta pc (43 – 52), hp3 much longer than hp2 (23 – 27). Palp length (195 – 212); apical margin of palptrochanter with a single spine anterolaterally; palptrochanter setae dissimilar in length, v1 48 – 52, v2 38 – 42.
Adult male. Gnathotectum as in female (Fig. 2C). Cheliceral shaft, excluding short basal section 105 – 130 long; fixed digit with offset subapical tooth followed by minute pilus dentilis inserted on small angular protrusion and row of 11-16 small teeth (Fig. 2H); movable digit (40 – 45) unidentate; spermatodactyl 45 – 54 long, 8 – 9 wide at medial region, extending anteriorly 28 – 37 beyond apex of digit, with narrowed neck and small bifid head apically (Figs. 2F, G). Corniculi more widely spaced but not more arched than in female, 33 – 37 long, and flanked medially by pair of short (7-10) spiculate processes (Figs 2D, 11A); internal malae and labrum as in female. Subcapitulum with seven rows of deutosternal denticles much as in female, sixth and seventh rows slightly widest; hypostomatic setae hp1 (28 – 32) similarly as long as pc (25 – 37), hp3 usually longest (30 – 50), hp2 (12 – 20). Palpi (length 145 – 187) similar in structure, form of setae as in female; palptrochanter with inner seta longer (30 – 44) than outer seta (17 – 32) but not markedly whip-like.
Legs — Adult female. Legs I (707 – 755) about as long as legs IV (715 – 750), and about as long as dorsal shield; other leg lengths (excluding pretarsi): II (565 – 600), III (587 – 615). Leg I pretarsus with biarticulate shaft (38 – 42 to base of claws), tarsus (187 – 205) about 1.6 longer than tibia; other length ratios, femur (135 – 147): genu (112 – 118): tibia (117 – 125) about 1.15: 0.95: 1.0. Legs II-IV with tarsus/tibia length ratios about 2.0-2.3. Chaetotactic formulae of leg segments as given for species-group. Tarsi II-IV with ventroapical process (12 – 17) bluntly pointed, smooth apically (Figs 13A, H), and with setae (d-1) (23 – 30) slightly shorter than pretarsus to base of claws (28 – 37). Tarsi II-IV with paradactyli asymmetrical, anterior element increasingly extending beyond tip of claw from II to IV, its length 30 – 35, twice as long as claw (15) on leg IV (Fig. 14A). Tarsi II and III with setae (l-1) and (v-1) symmetrical in form, pl-1 (32 – 35) and av-1 (36 – 47) slightly longer than al-1 (27 – 30) and pv-1 (26 – 37), respectively. Tarsus IV with distal seta av-1 (50 – 60) blunt, clearly larger, thicker than pv-1 (37 – 45); adjacent setae (l-1) symmetrically short (20 – 27), slender (Fig. 14A). Tarsi II-IV with dorsal seta md nearly reaching base of claws, and nearly as long as smooth attenuated setae (d-2) (67 – 80 on II, 85 – 100 on III, 95 – 110 on IV).
Adult male. Legs I clearly longer than dorsal shield, not thicker than those of female; relative leg lengths, excluding pretarsi, similar to those of female, I 550 – 660, II 445 – 524, III 455 – 552, IV 575 – 675. Legs with segment length ratios as in female. Tarsus I with long pretarsus (25 – 37 to base of claws), claws 10 – 12 long. Tarsi II-IV with dorsoapical setal processes (d-1), ventroapical process and pretarsal structures formed as in female, pretarsus lengths 27 – 35 excluding claws (10) (Fig. 13B). Legs III-IV with tibial setae (v) and leg IV genual seta av dimorphically thicker, spinelike, less attenuate; length and form of other setae much as in female; seta pl-3 of basitarsus II smooth, attenuate (36 – 38) (Fig. 13B), but pl-3 of basitarsi III, IV (21 – 22) similar to pl-2 in thicker, slightly shorter form than other three basitarsal setae.
Type material — Except the final one, noted below, all specimens collected at the La Selva Biological Station, Heredia Province, Costa Rica (10°26´N, 84°01´W, elevation 50-150 m): HOLOTYPE: adult female, 8 July 1997, ex adult Cephaloleia bella Baly on Heliconia pogonantha leaves, coll. M. Paniagua. PARATYPES: 1 adult female, 1 adult male, with same data as holotype; 2 adult females, 1 adult male, 12 June 1997, ex coexistent adults of Cephaloleia belti Baly and C. ornatrix Donckier from Heliconia leaves, coll. ALAS; 3 adult females, 2 adult males, 15 April 1994, ex adult Chrysomelidae, no host plant data, M. Paniagua coll.; 1 adult female, 1 adult male, June 1997, ex adult Cephaloleia bella Baly, no host plant data, coll. D. Johnson; 1 adult female (gravid with larva), 1 adult male, June 1997, ex adult Cephaloleia belti Baly, no host plant data, coll. D. Johnson; 2 adult males, February 1994, associated with adult Chelobasis sp. on leaves of Heliconia pogonantha, coll. E.E. Lindquist & ALAS; 1 adult female, 2 adult males, 18 February 1994, ex adult Chrysomelidae, no host plant data, coll. ALAS; 3 adult females, 2 June 1997, with adult hispine beetles on Heliconiaceae leaves, coll. E.E. Lindquist; 2 adult females, June 1997, ex adult Cephaloleia vicina on Heliconia pogonantha, coll. not indicated; 1 adult male, February 1994, associated with adult Cephaloleia vicina on leaf of Heliconia pogonantha, coll. E.E. Lindquist & ALAS. Costa Rica, Osa Peninsula, Corcovado National Park (8°27.39´N, 83°25.45´W,elevation 10-500 m): 1 adult male, May 1997, ex adult Chelobasis bicolor Gray, no host plant data, coll. D. Johnson.
Host identifications — Some of the identities of the associated beetles given above are in accord with current molecular studies, i.e., those as noted below for Lasioseius serripes. However, the following identities are discordant. Cephaloleia vicina does not occur at La Selva, and specimens identified as such from there are, in all cases checked by DNA barcodes, C. bella. Further, C. bella normally does not occur on Heliconia pogonantha; thousands of field diet records indicate it feeds only on H. imbricata (Garcia-Robledo et al., 2017).
Etymology — The specific name is based on the chrysomelid subfamilial name Cassidinae, which includes the tribes Arescini and Imatidiini (or Cephaloleini) to which beetle associates of the genera Chelobasis and Cephaloleia belong, respectively. We have chosen this name for the species with the widest array of host associates and as the species-group name, anticipating that rolled-leaf beetles of other genera in these tribes will be found to live with these mites.
Remarks — The male from Corcovado measured ca 12-19% smaller in dimensions than seven males measured from La Selva; however, in other descriptive aspects it fully resembles those from La Selva. Two females are asymmetrical in having humeral seta r3 inserted off the dorsal shield on one side, yet the aligned r2 and r4-r6 are on the shield. Two females show asymmetry in size and arrangement of the vertical cluster of setae j1, j2, z1; one male lacks one j2.
Collection records of this mite species associated with beetles of three different species of Cephaloleia and one of Chelobasis leave uncertainty as to host preferences, particularly as beetles of these species are often coexistent (Garcia-Robledo 2010, Staines 2011, Schmitt & Frank 2013, Moraza & Lindquist 2015). Adults of Lasioseius cassidini are clearly the largest of the described species of this group. However, the data available are inadequate to test whether a mite/beetle size correlation exists, such as was hypothesized for larger mites being associated with larger beetles, for species of Hispiniphis and Makarovaia, which are more intimate subelytral associates of these same beetles (Moraza & Lindquist 2015, 2016). Although there are records of a few specimens of L. cassidini with relatively large beetles of Chelobasis, including one from the Osa Peninsula, they are based only on adult male mites, which may not present inherent preferences phoretically. Tentatively, subject to further studies, we conclude that, at La Selva, Lasioseius cassidini may be associated regularly with small beetles of the host specialist Cephaloleia bella and to some extent with the host generalist C. belti on Heliconia imbricata.
ZOOBANK: F1A4EE0C-0031-410D-B993-20CDC3D08D71 ![]()










Diagnosis — Adult female: Sternal shield reticulated over entire surface, its posterolateral corners emarginated lateral to setae st3; poroids iv3 and setae st4 on soft cuticle; epigynal shield fully reticulated, its lateral margins parallel anterior to setae st5; ventrianal shield with posterolateral margins on either side of anus divergent, and with three pairs of opisthogastric setae, JV1 on soft cuticle; metapodal plates connected or coalesced on each side. Dorsal setae r6 usually on soft cuticle; Z5, S5 attenuated like adjacent setae. Adult male: Sternitigenital shield reticulated over entire surface. Leg II with dorsal setae dimorphically short, blunt on genu, tibia, tarsus. Tibiae III-IV with seta pv dimorphically hooked, blunt. Adult attributes: Gnathotectum anterior margin truncate, irregularly denticulate. Fixed chela with file of ca 20 teeth in female, ca 18 in male. Sternal setae st1 inserted on anterior edge of sternal shield. Tarsi II-IV with ventroapical process truncate-spiculate apically; tarsi II-III with seta av-1 slightly longer and thicker at base than pv-1and both slightly longer than (l-1); tarsus IV with apical setae (v-1) symmetrically thicker than, and about twice as long as (d-1); pretarsal paradactyli small, symmetrical.
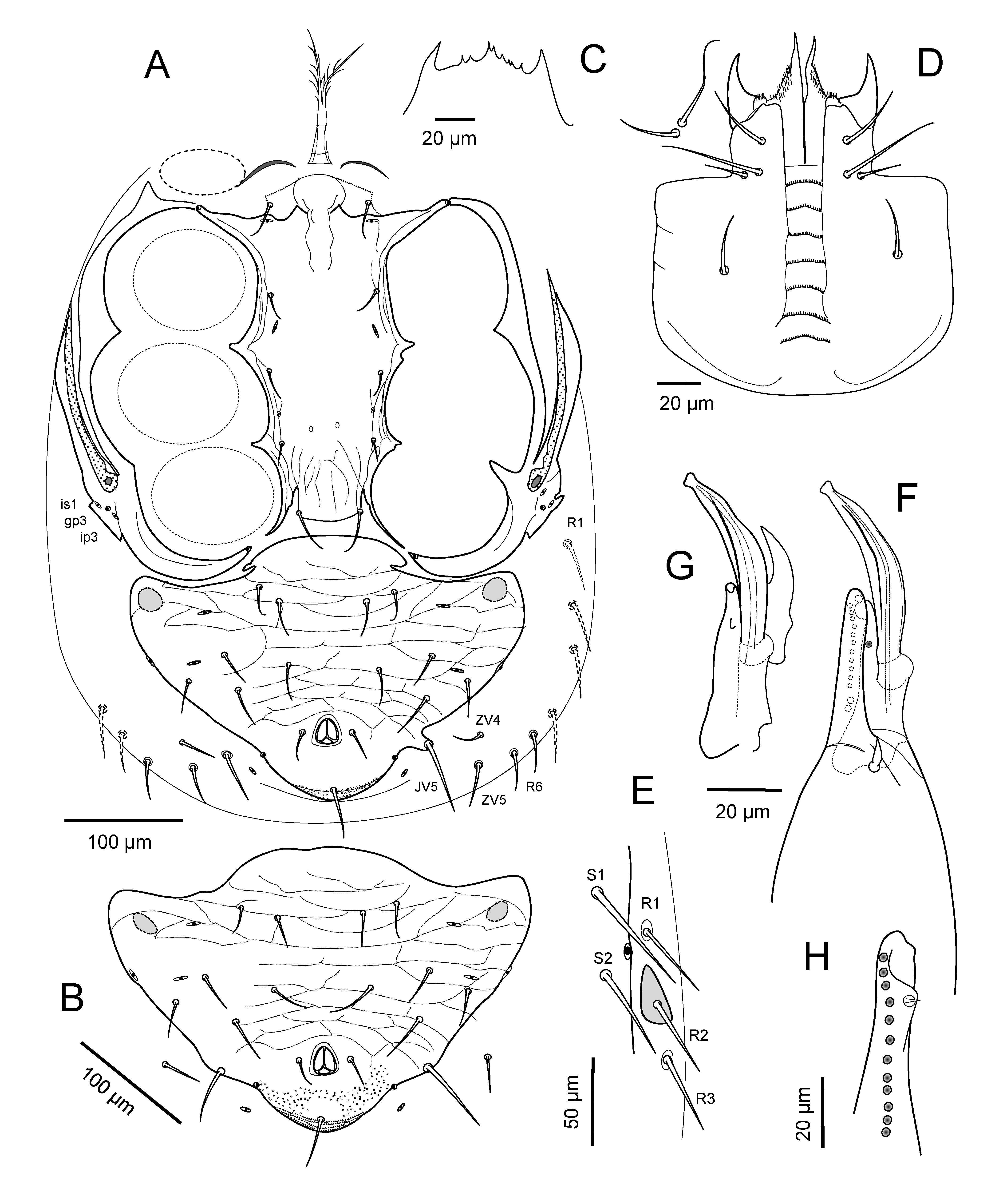
Idiosomatic dorsum — Adult female: Dorsal shield 651 – 695 long, 337 – 385 wide at level of r3, 368 – 397 wide at level of r5 (n=9), lightly reticulated over entire surface, more faintly so behind setae Z4, where also faintly punctate. Shield with 37 pairs of setae, all acutely pointed (Figs. 3A, B), with few barbs, except J5 short (15 – 17), clearly barbed; J1-J4 about as long as longitudinal intervals between their bases, Z1-Z4 and S2-S4 longer than intervals between their bases; r3 erect, similarly as long as r2, r4: j1 (41 – 50), j2 (45 – 55), j3 (60 – 70), j4 (43 – 50), j5 (48 – 55), j6 (58 – 63), z1 (20 – 29), z2 (50 – 55), z3 (52 – 60), z4 (64 – 70), z5 (47 – 55), z6 (60 – 70), s1 (48 – 55), s2 (50 – 60), s3 (53 – 65), s4 (64 – 70), s5 (63 – 75), s6 (57 – 72), r2 (50 – 60), r3 (54 – 62), r4 (55 – 62), r5 (56 – 67), J1 (45 – 55), J2-J4 (48 – 62), Z1-Z3 (71 – 97), Z4 (87 – 102), Z5 (70 – 88), S1(57 – 70), S2 (66 – 77), S3 (64 – 75), S4 (66 – 78), S5 (64 – 75). Lateral soft cuticle with eight pairs of smooth setae, including r6 (52 – 62), R1 (48 – 55), R2-R6 (44 – 52) and ZV5 (43 – 47) posteriorly.
Adult male. Dorsal shield 557 – 587 long, 332 – 357 wide at level of setae r3, 325 – 353 wide at level of r6 (n = 5); shield ornamentation, complement of pore-like structures and setation as in female except slightly more expanded laterally so as to become united with peritrematal shields at level of setae s2 and bear marginal setae r6 and sometimes R1 (Fig. 5A). Dorsal shield with 37-38 pairs of setae, their form and relative lengths as in female, except bases of Z4, Z5, S4, S5 swollen sub-basally. Setae j1 (28 – 33), j2 (40 – 43), j3 (53 – 60), j4 (38 – 40), j5 (38 – 47), j6 (48 – 54), z1 (17 – 20), z2, z3 (45 – 50), z4 (55 – 60), z5 (38 – 44), z6 (50), s1 (46 – 48), s2 (52 – 56), s3 (50 – 61), s4 (58 – 65), s5 (58 – 65), s6 (55 – 58), r2-r4 (48 – 53), r5 (57 – 60), r6 (50 – 52), J1-J4 (40 – 47), J5 (9 – 12), Z1-Z3 (66 – 75), Z5 (61 – 72), S1(50 – 52), S2 (55 – 57), S3-S5 (57 – 66), Z4 the longest opisthonotal setae (75 – 86). Lateral soft cuticle with usually six pairs of marginal setae (40 – 47). Form of peritrematal shields as in female.
Idiosomatic venter — Adult female. Tritosternum base 28 – 33 long, 16 – 20 wide at its base, with paired laciniae fused along basal 15 – 22 of entire length (95 – 115). Presternal region with circumscribed pair of weakly lineated granulated areas adhered to anterior margin of sternal shield. Sternal shield 100 – 112 long, with irregularly concave posterior margin, 118 – 125 wide, well reticulated over entire surface, midsurface with narrower reticula; sternal setae of similar length, st1 (38 – 42), st2 (33 – 39), st3 (35 – 42). Sternal poroids iv3 and setae st4 (26 – 32) inserted on soft cuticle. Endopodal strips narrow between coxae III and IV. Epigynal shield well reticulated over entire surface, with narrower reticula anterior to insertion of st5 (36 – 42), its broadly rounded hyaline anterior margin not reaching posterior edge of sternal shield, its lateral margins nearly parallel anterior to setae st5, and its posterolateral corners slightly to clearly emarginated behind st5; shield 188 – 210 long, 95 – 107 at widest level at posterior angulate margin, 62 – 70 at narrowest width between legs IV. Opisthosomatic venter with two pairs of metapodal platelets connected or coalesced on each side, primary pair ovate, 25 – 30 long, 13 – 17 wide, secondary pair with greatest dimension transverse, 13 – 22; one adjacent set of small, narrow platelets near anterior margin of ventrianal shield, lateral to JV1. Ventrianal shield inversely subtriangular with rounded anterior corners, with posterolateral margins on either side of anus divergent anteriorly; surface strongly reticulate except faintly in anal region where also faintly punctate on either side of postanal seta; shield wider (253 – 295) than long (179 – 205), including cribrum (8 – 12); shield with three pairs of ventral setae and sometimes pores gv3: JV2 (33 – 40), JV3(35 – 45), ZV2 (34 – 40); with paranal setae (20 – 27) shorter than postanal seta (49 – 62). Soft cuticle with six pairs of simple opisthogastric setae, JV1 and ZV1 (26 – 33) anterior to ventrianal shield, ZV3 (32 – 40), ZV4 (40 – 47), JV5 (55 – 65) flanking the shield, and ZV5 (43 – 47) aligned behind R6; three pairs of poroids (including ivp) and sometimes gland pore gv3 on soft opisthogastric cuticle. Peritrematal-exopodal shield with lines extending from stigma to irregularly rounded posterior margin (Fig. 4A); peritreme extending to level slightly anterior to insertion of setae s1 (Fig. 3A). Spermathecal apparatus with cup-like calyx ca. 30 – 35 long from atrium at base, 23 – 26 wide at distal margin (Fig. 3D).
Adult male. Tritosternum much as in female, but with base biarticulate, longer (21 – 24) than wide (13 – 17), and laciniae (78 – 88) fused along basal 10 – 12 of entire length (Fig. 5C). Presternal region weakly granulated, with or without pair of weakly sclerotized transverse strips anteriorly. Sternitigenital shield length 245 – 283 from anterior margin of genital opening, widths 104 – 110 at level between coxae II, 92 – 100 between coxae III, and 58 – 71 between coxae IV; shield nearly fully reticulated, but bare near posterior margin where endopodal apices freely truncated beside coxae IV; shield medial region more weakly ornamented but punctate between setae iv1 and st3, and with poroids iv3 appearing as distinctively rounded pits; setae st1 on anterior margin of sclerotized region of shield; setae st1-st5 of dissimilar lengths as in female, st1-st3 (27 – 30), st4 shortest (20 – 23) and st5 longest (30 – 35). Ventrianal shield strongly reticulate as in female, its greatest width (282 – 305) larger than its length (178 – 220, including cribrum ca. 9 – 11 long, 75 – 86 wide); shield with seven pairs of opisthogastric setae; lateral margins irregular or eroded near where setae ZV4 on adjacent soft cuticle; setae JV1 (30 – 32), JV2 (37 – 42), JV3 (37 – 44), ZV1 (27 – 30), ZV2 (35 – 38), ZV3 (28 – 33), ZV4 (35 – 40), and JV5 the longest (50 – 59) inserted in bluntly angled lateral corners; postanal seta smooth (40 – 48), about twice as long as paranal setae (19 – 25). Soft opisthogastric cuticle with two pairs of setae, including ZV5 aligned behind R6, and one pair of lyrifissures, ivp (Fig. 5E).
Gnathosoma — Adult female. Gnathotectum anterior margin broadly truncate, variably with two lateral, short, smooth prongs or with few denticles, sometimes with smaller denticles between lateral prongs (Fig. 3C). Cheliceral shaft, excluding basal section, 136 – 148 long, with slender digits (Figs 3E, F); dorsal seta blunt (8 – 11); fixed digit with tiny offset subapical tooth and row of 19-24 very fine teeth (Fig. 3E), pilus dentilus minute (1 – 2) at level of 4th apical tooth; movable digit (62 – 72) tridentate. Subcapitulum rows of deutosternal denticles each with numerous (ca 25-40) fine denticles, those of most anterior row slightly coarser (ca 25-30), 2nd-5th rows with ca 35-40 teeth, 6th and sometimes 7th rows slightly wider and slightly more denticulate (Fig. 4D); subcapitular setae hp1 (30 – 37) slightly shorter than capitular setae pc (36 – 42), hp3 slightly longest (40 – 47), much longer than hp2 (23 – 27). Salivary styli and corniculi as in figure 4C. Palp length (193 – 202); apical margin of palptrochanter without spines anterolaterally; palptrochanter setae dissimilar in length, v1 45 – 63 and v2 34 – 42 (Figs 4E, F).
Adult male. Gnathotectum as in female (Fig. 5C). Cheliceral shaft, excluding short basal section, 110 – 120 long (Fig. 5F); fixed digit with offset subapical tooth followed by pilus dentilis behind hollow area and row of 10-16 small-sized teeth (Figs. 5F, G); movable digit (42 – 45) unidentate; spermatodactyl 40 – 57 long, 6 – 7 wide at base, slightly wider (8) at medial region, extending anteriorly ca 35 beyond apex of digit, with narrowed neck and small head apically (Figs 5F, H). Corniculi slightly more widely spaced and arched inwardly than in female, 25 – 30 long, and flanked medially by pair of short (7 – 8) spiculate processes (Figs 5I, 11B); internal malae and labrum as in female. Subcapitulum with seven or rarely eight rows of deutosternal denticles formed much as in female; hypostomatic setae hp1 not attenuated (21 – 27), slightly shorter than capitular setae pc (22 – 32), hp3 (30 – 37) slightly longest, hp2 shortest (15 – 20) (Fig. 5I). Palpi (length 173 – 188) similar in structure, form of setae as in female; palptrochanter with inner seta longer (38 – 40) than outer seta (20 – 23) but not markedly whip-like (Fig. 5I).
Legs — Adult female. Legs I (669 – 715) nearly as long as legs IV (678 – 725), and slightly longer than dorsal shield; other leg lengths (excluding pretarsi): II 519 – 529, III 538 – 545 (Figs 6A-E). Leg I pretarsus with biarticulated shaft (38 – 45 to base of claws), claws 15 – 17 long; tarsus (173 – 182) about 1.4-1.5 longer than tibia; other length ratios, femur (137 – 145): genu (110 – 117): tibia (112 – 124) about 1.2: 0.95: 1.0. Legs II-IV with tarsus/tibia length ratio about 2.0-2.2 (Figs 6C, D). Chaetotactic formulae of leg segments as given for species-group. Tarsi II-IV with ventroapical process (15 – 17) truncate-spiculate apically (Figs 13C, 14B), and with setae (d-1) (25 – 30) shorter than pretarsus to base of claws (33 – 44). Tarsi II-IV with paradactyli nearly symmetrical, weakly developed, anterior element barely extending beyond tip of claw (Figs 13G, 14B). Tarsi II (Fig. 13C), III with setae (l-1) (44 – 48) shorter than (v-l) (50 – 65); dorsal seta md (77 – 85) longest of tarsal setae, (d-2) barbed, less attenuated (50 – 62). Tarsus IV with setae av-1 (70 – 75) slightly longer than pv-1 (60 – 65), both nearly twice as long as similarly slender (l-1) (38 – 42) (Fig. 14B).
Adult male. Legs I slightly thicker than those of female, and longer than dorsal shield; relative lengths similar to those of female, I 690 – 705, II 475 – 525, III 493 – 507, IV longest, 710 – 735. Legs with segment length ratios as in female. Tarsus I with long pretarsus (25 – 35 to base of claws), claws 15 long. Tarsi II-IV with spiculated form of ventroapical process (Fig. 13D), relative lengths of dorsoapical setal processes (d-1) (25 – 28) and pretarsal structures as in female; tarsi II-III with setae (v-1) (45 – 55) symmetrically longer than slender (l-1) (33 – 44), increasingly so on III; tarsus IV with av-1 (70 – 75) and pv-1 (55 – 67) asymmetrically longer than (l-1) (32 – 35) (Fig. 13D). Legs II with dorsal setae of genu, tibia, basitarsus and (d-2) of telotarsus (16, 23, 26, 27), distinctively shorter than in female and bluntly spine-like (Figs 12A, 13D); genu III and tibiae III-IV seta pv and tibia IV seta av dimorphically hooked, blunt (Fig. 12B). Length and form of other setae slightly thicker than in female.
Deutonymph. None available to study for comparative purposes.
Protonymph. General description, based on one specimen, without measurements, for species-group perspective. Dorsal idiosoma (Fig. 7C) with setae generally nearly aciculate, but with some setae acquiring slightly tricarinate or barbed form as in adult. Podonotal region holotrichous, with 15 pairs of setae (five pairs added to larval complement, including j2, s5 amidst 11 pairs on podonotal shield and r2, r3, r5 along with larval s6 on lateral soft cuticle); j2 inserted in nearly transverse alignment with j1. Opisthonotal region holotrichous, with 15 pairs of setae (five pairs added to larval complement, including J1, Z1, Z2, S2, R1, all on interscutal soft cuticle; pygidial shield expanded so as to include eight pairs of setae, including J3, Z5, S4, S5 (all larval setae).
Ventral idiosoma (Fig. 7D) with all setae aciculate, with no postlarval podosomal additions except st5. Sternal poroids iv1, iv2 present, iv3 absent, and larval funicules suppressed. Anal shield oval, with paranal setae similar in length or slightly shorter than postanal seta; rudiments of euanal setae suppressed. Opisthogastric soft cuticle with four or five pairs of setae, with asymmetrical addition of ZV5, which usually deutonymphal; JV5 relatively larger than in larva, and inserted more posteriorly, in transverse alignment with postanal seta.
Gnathosoma — Fixed cheliceral digit with even row of 12-13 teeth; movable digit tridentate. Gnathotectum similar in form to adult. Subcapitulum with four pairs of setae, hypostomatic h1-h3 and capitular pc. Palpus with normal protonymphal complement of setae, including one on trochanter (Lindquist & Evans 1965); palptarsus with hyaline flap developed beside apotele as in adult.
Legs — Legs I-IV with claws, pretarsi and shape of setae similar to adult female; tarsi II-IV with ventroapical process similar in form, serrated apically, as adult. Legs I, II, III, IV with normal, holotrichous protonymphal chaetotaxy (Lindquist & Evans 1965), except genu IV with accelerated presence of usually deutonymphal seta pl-1, and femur IV with accelerated alveolar indication of seta v-1 near basal lyriform fissure: coxae 2, 2, 2, 1; trochanters 4, 4, 4, 4; femora 10 (2 2/1 2/2 1), 8 (1 2/1 2/1 1), 5 (1 2/1 1/0 0), 4 (1 2/0 1/0 0); genua 8 (1 2/1 2/1 1), 6 (1 2/0 2/0 1), 6 (1 2/0 2/0 1), 6 (1 2/0 2/0 1); tibiae 8 (1 2/1 2/1 1), 7 (1 1/1 2/1 1), 7 (1 1/1 2/1 1), 7 (1 1/1 2/1 1); tarsi II-IV each with 17 (md added to larval complement). Tarsus IV with seta av-1 distinctively erect, longer and slightly thicker than all other tarsal setae (Fig. 12G), in contrast to its form on tarsi II, III of protonymph, and on tarsus IV of adult.
Larva. General description, based on two unborn specimens, without measurements, for species-group perspective. Dorsal idiosoma (Fig. 7A) with all setae aciculate. Podonotal shield holotrichous with typical nine pairs of setae (j1, j3-j6, z2, z4, z5, s4); setae s6 on soft cuticle behind posterolateral margin of shield. Opisthonotal region holotrichous with 10 pairs of setae ( J2-J5, Z3-Z5, S3-S5); pygidial shield margins mostly indiscernible on specimens at hand, but its surface with typical pair of arcuate lines near insertions of setae J4, and with typical complement of four pairs of setae, including tiny J4, J5 and prominent Z3, Z4. Soft interscutal cuticle with two pairs of mesonotal scutellae and typical six pairs of setae, including moderately long J2, J3, S3, S5, prominent Z5 (in ventrolateral position), and shorter S4.
Ventral idiosoma (Fig. 7B) with all setae aciculate. Sternal shield outline not distinguishable; setae st1-st3 similarly short, st4, st5 absent, but the typical pair of subcutaneous structures (‘funicules’ sensu Athias-Henriot 1971a) present posteromediad bases of legs III. Anal shield margins indistinguishable; anal valves with rudimentary pair of euanal setae; paranal setae slightly longer than postanal seta. Opisthogastric soft cuticle with typical four pairs of similarly short setae ( JV1, JV2, JV5, ZV2).
Gnathosoma — Fixed cheliceral digit with even row of 10-11 teeth; movable digit bidentate. Gnathotectum triramous, branches shorter than in adult. Subcapitulum with two pairs of hypostomatic setae, h1 longer than h2. Palpus with normal larval complement of setae (Lindquist & Evans 1965); palptarsus with hyaline flap not evident beside apotele.
Legs — Legs I-III with claws, pretarsi and shape of setae similar to adult female; tarsi II, III with ventroapical process similar in form, serrated apically, as adult, and with setae (d-1) more strongly developed, slightly longer than pretarsi to base of claws. Legs I, II, III with normal, holotrichous larval chaetotaxy (Lindquist & Evans 1965): trochanter 2, 2, 2; femora 10, 7, 5; genua 8, 6, 6; tibia 8, 7, 7; tarsi II, III each with 16.
Type material — All specimens collected at the La Selva Biological Station, Heredia Province, Costa Rica (10°26´N, 84°01´W, elevation 50-150 m): HOLOTYPE: adult female, 12 June 1997, ex Heliconia leaves, successional plots, coll. ALAS. PARATYPES: 5 adult females, 5 adult males, 1 protonymph, with same data as holotype; 3 adult females (2 gravid with larva), 6 adult males, 12 June 1997, ex coexistent adults of Cephaloleia belti Baly and C. ornatrix Donckier from Heliconia leaves, coll. ALAS; 1 adult female (CRI 002: 739027), 8 July 1997, ex adult Chelobasis perplexa Baly, no host plant data, coll. M. Paniagua; 3 adult females, 1 adult male, June 1997, ex adult Cephaloleia ornatrix, no host plant data, coll. D. Johnson.
Host identifications — The identities of the associated beetles given above are in accord with current molecular studies, which indicate that one haplotype of Chelobasis perplexa sensu stricto and records of Cephololeia belti and C. ornatrix on Heliconia pogonantha are in accord with field diet records. While C. belti is a host generalist, C. ornatrix is a host specialist of Heliconia pogonantha (Staines & Garcia-Robledo 2014, Garcia-Robledo et al. 2017).
Etymology — The specific name is a Latinized adjectival combination of ‘serra’, meaning saw or toothed like a saw, and ‘pes’, meaning foot, and refers to the unusual apically serrated form of the ventral apical tarsal process of legs II to IV.
Remarks — One female is asymmetrical in having seta r6 on edge of dorsal shield on one side, rather than on soft cuticle. As noted under morphological remarks following description of the cassidini species-group, the rudimentary presence of seta av-1 on femur IV and full presence of seta pl-1 on genu IV are ontogenetic accelerations of setae that are typically first expressed in the deutonymph (Evans 1963).
Although far from definite, collection records of Lasioseius serripes indicate a frequent association with Cephaloleia ornatrix, which perhaps is the preferred companion. Tentatively, subject to further studies, we conclude that, at La Selva, Lasioseius serripes is associated regularly with the host specialist C. ornatrix and to some extent with a form of Chelobasis and the host generalist C. belti on H. pogonantha.
ZOOBANK: F088E970-F8B8-4E76-8382-87DA2EF12B7F ![]()



Diagnosis — Adult female: Sternal shield lineated laterally and anteromedially, its anterior margin with blunt medial projection, its posterolateral corners not emarginated; poroids iv3 and setae st4 on slender metasternal platelets; epigynal shield reticulated over nearly entire surface, its lateral margins concave, narrowed anterior to setae st5; ventrianal shield with posterolateral margins on either side of anus nearly parallel, and with four pairs of opisthogastric setae, JV1 on shield; metapodal plates nearly connected or coalesced on each side. Dorsal setae r6 on edge of dorsal shield; Z5, S5 attenuated like adjacent setae. Adult male: Sternitigenital shield incompletely reticulated, nearly smooth medially. Dimorphic leg setal structures minimal. Adult attributes: Gnathotectum anterior margin with 3 elongated, denticulated tines. Fixed chela with file of 8-9 teeth in female, ca 4 in male. Sternal setae st1 inserted on anterior edge of sternal shield. Tarsi II-IV with ventral apical process bluntly pointed, smooth apically; tarsi II-III with setae (l-1) shorter, more spinelike than (v-1); tarsus IV with distal seta pv-1 longer, more attenuate than av-1, (l-1); pretarsus with anterior paradactylus slightly longer than posterior one.
Idiosomatic dorsum — Adult female. Dorsal shield 424 – 450 long, 201 – 245 wide at level of r3, 267 – 296 wide at level of r6 (n=7), lightly reticulate-lineate on lateral surfaces but nearly smooth medially between bases of setae j4 to J4, and sparsely punctate behind level of setae Z4; shield with 38 pairs of setae generally similar in size and acutely pointed form, with few apical barbs, more so on Z4, Z5, S4, S5 (Figs 8A, B); J1-J3 and Z1-Z4 slightly longer than longitudinal intervals between their bases, S2-S4 clearly so; J5 minute, barbed (4 – 6): j1 (28 – 31), j2 (26 – 32), j3 (41 – 44), j4 (26 – 32), j5 (32 – 35), j6 (45 – 47), z1 (19 – 24), z2 (30 – 35), z3 (37 – 42), z4 (41 – 45), z5 (32 – 37), z6 (41 – 46); s1 (32 – 38), s2 (34 – 43), s3 (40 – 45), s4 (43 – 48), s5 (45 – 50), s6 (47 – 52), r2 (35 – 42), r3 erect (21 – 30), r4 (32 – 48), r5 (32 – 50), r6 (32 – 48), J1 (32 – 42), J2 (40 – 50), J3 (43 – 52), J4 (43 – 52), Z1 (48 – 51), Z2 (51 – 54), Z3 (54 – 58), Z4 (58 – 62), Z5 (63 – 70), S1 (43 – 50), S2 (44 – 53), S3 (46 – 58), S4 (46 – 58), S5 (43 – 52). Lateral soft cuticle with seven pairs of smooth setae, including R1 (40 – 48), R2-R6 (35 – 42) and ZV5 posteriorly (30 – 33) (Fig. 8D). Peritrematal shield fused to dorsal shield at level of s1.
Adult male. Dorsal shield 338 – 340 long, 185 – 200 wide at level of setae r3, 200 – 215 wide at level of r6 (n = 2); shield ornamentation, complement of pore-like structures and setation as in female except slightly more expanded laterally so as to become united with peritrematal shields at level of setae s2 and bear setae R1. Dorsal shield with 39 pairs of setae, their attenuated form and relative lengths as in female. Setae j1 (21 – 23), j2 (16 – 26), j3 (35 – 36), j4 (21 – 25), j5 (24 – 25), j6 (32 – 33), z1 (14 – 16), z2 (22), z3 (32), z4 (34 – 35), z5 (27 – 28), z6 (32 – 36), s1 (29 – 30), s2 (30 – 31), s3 (34 – 38), s4 (36), s5 (42), s6 (35 – 36), r2, r4, r5 (33 – 37), r3 erect, shorter (23 – 27), r6 (36 – 38), J1-J3 (27 – 32), J4 (25 – 27), J5 (3 – 4), Z1-Z2 (40 – 43), Z3 (44 – 45), Z4, Z5 longest (48 – 54), S1, S2 (35 – 41), S3 (41 – 45), S4 (40 – 43), S5 (32 – 35). Lateral soft cuticle with six pairs of setae, R2-R6 (27 – 37) followed by ZV5.
Idiosomatic venter — Adult female. Tritosternum base 26 – 28 long and 10 – 14 wide at its base, with paired laciniae fused for about 33 – 37 of entire length (96 – 100). Presternal region with circumscribed pair of jagged-lineate, granulated areas adhered to anterior margin of sternal shield, sometimes flanked anteriorly by a pair of narrow slightly sclerotized strips on either side of tritosternum base. Sternal shield 87 – 93 long from slight, blunt medial projection of anterior margin to gently concave posterior margin, 85 – 95 wide, weakly reticulate-lineate over much of surface or nearly smooth medially, with arcuate transverse line between poroids iv1 (Fig. 8D); sternal shield setae of similar lengths, st1 (15 – 20), st2, st3 (11 – 16). Sternal poroids iv3 and setae st4 (6 – 14) inserted together on small metasternal platelets (15 – 20). Endopodal strips narrow between coxae III and IV. Epigynal shield lightly reticulate over entire surface, its narrowly rounded hyaline margin nearly reaching or slightly overlapping posterior edge of sternal shield, its lateral margins concave, narrowed anterior to setae st5 (11 – 15), and its posterolateral corners slightly emarginated behind st5; shield 140 – 147 long, 78 – 87 at widest level at posterior rounded corners, 66 – 71 at level of st5, and 49 – 54 at narrowest width between legs IV. Opisthosomatic venter with two pairs of metapodal platelets touching or coalesced on each side, primary pair ovoid to subtriangular, 20 – 25 long, 10 – 12 wide, secondary pair smaller, greatest transverse dimension 8 – 12. Ventrianal shield inversely subtriangular with irregularly rounded anterior corners, with posterolateral margins on either side of anus nearly parallel; surface entirely lightly sculptured, with primarily transverse lineation changing to reticulation behind setae JV2-ZV2, except anal region sparsely punctate on either side of postanal seta; shield wider (160 – 183) than long including cribrum (136 – 147), width at anal level (95 – 104); gland pore gv3 on shield edge posterior to anal opening; shield with four pairs of ventral setae: JV1 (12 – 17), JV2 (22 – 27), JV3 (24 – 31), ZV2 (22 – 27); with paranal setae (11 – 14) shorter than postanal seta (28 – 35). Soft cuticle with four pairs of simple opisthogastric setae, ZV1 (11 – 15) anterior to ventrianal shield, ZV3 (17 – 22), JV5 attenuated, barbed (48 – 55) flanking the shield, and ZV5 (30 – 33) somewhat aligned behind R6; three pairs of poroids (including ivp) on soft opisthogastric cuticle, ivo2 sometimes captured on anterolateral edge of ventrianal shield. Peritrematal-exopodal shield with lines extending from stigma into rounded posterior margin (Fig. 8D); peritreme extending to level between setae s1 and z1 (Fig. 8A). Spermathecal apparatus with cup-like calyx (12 – 15 long including embolus).
Adult male. Tritosternum much as in female, but with base biarticulate, wider (6 – 11) than long (5 – 10), and laciniae (37 – 40) fused (8) for only about 1/5 of total length. Presternal region weakly ornamented, sometimes with pair of narrow strips on either side of tritosternum base. Sternitigenital shield length 158 – 162 from genital opening, widths 65 – 72 at level between coxae II, 50 between coxae IV; shield with a concave arcuate line between poroids iv1, lightly lineate-reticulate along lateral margins and posteriorly between coxae IV, unornamented medially between st2 and iv3, with endopodal apices freely acuminate beside coxae IV; setae st1-st5 of dissimilar lengths as in female, st1 (14 – 16), st2 (11 – 12), st3 (8 – 11), st4 shorter (4 – 8), st5 (7 – 8). Ventrianal shield reticulate as in female, its convex anterolateral margins entire behind apices of exopodal strips behind coxae IV, and posterolateral margins not constricted in anal region, its greatest width (173 – 189) larger than its length (123 – 141, including cribrum ca. 3 – 8 long, 73 – 80 wide); shield with seven pairs of opisthogastric setae, JV1, ZV1 (10 – 12), JV2, JV3 (25 – 29), ZV2 (20 – 26), ZV3 (17 – 20), and longest JV5 (40 – 43); postanal seta smooth (26 – 28), about three times as long as paranal setae (8 – 12) (Fig. 9A). Soft opisthogastric cuticle with two pairs of lyrifissures including ivp.
Gnathosoma — Adult female. Gnathotectum anterior margin with three denticulate prongs of similar moderately elongate length or medial prong longer than laterals (Fig. 8C). Cheliceral shaft, excluding basal section, 139 – 155 long, with slender digits; dorsal seta simple (10 – 11); fixed digit with tiny subapical tooth and row of 7-10 fine teeth, pilus dentilis minute, at level of 3rd apical tooth; movable digit (48 – 51) tridentate (Fig. 8E). Subcapitulum with deutosternal rows differing little in width, most anterior row with 9-10 teeth, 2nd to 5th rows with 8-15 teeth, basalmost two rows slightly wider, with ca 15-20 teeth (Fig. 8F); subcapitular setae hp1 (40 – 50) about as long as capitular setae pc (40 – 46), longer than hp3 (27 – 29), and hp2 (20 – 22). Palp length (113 – 122), apical margin of palptrochanter with two or usually three spines anterolaterally; palptrochanter with internal seta v1 sickle-shaped, longer (35 – 44) and thicker than v2 (24 – 30) (Fig. 8G).
Adult male. Gnathotectum anterior margin with elongated prongs as in female (Fig. 9B). Cheliceral shaft, excluding short basal section 91 – 98 long; fixed digit with offset subapical tooth followed by pilus dentilis and sparse row of 4-5 small teeth (Fig. 9E); movable digit (27 – 28) unidentate; spermatodactyl 30 – 40 long, 3 – 6 wide at base, extending anteriorly ca 23 – 30 beyond apex of digit, with narrowed neck and small head apically (Fig. 9D). Corniculi slightly more widely spaced but not more arched than in female, 24 – 26 long, their bases flanked medially by pair of short (5 – 7) spiculate processes (Figs 9C, 11C); internal malae and labrum as in female. Subcapitulum with rows of deutosternal denticles much as in female, anterior five rows with ca 8-15 teeth, 6th and 7th rows with ca 15-20 teeth; hypostomatic setae hp1 (28 – 30) slightly attenuated, about as long as pc (24 – 31), longer than hp3 (20 – 21), and hp2 shortest (10 – 12) (Fig. 9D). Palpi (length 96 – 101) similar in structure, form of setae as in female; apical margin of palptrochanter with three spines anterolaterally; palptrochanter with inner seta (26 – 30) longer than outer seta (8 – 11) (Fig. 9D).
Legs — Adult female. Legs I (357 – 385) subequally as long as legs IV (360 – 380), and shorter than dorsal shield; other leg lengths (excluding pretarsi): II 286 – 319, III 288 – 303. Leg I pretarsus with biarticulated shaft (16 – 21 to base of claws), claws 10 – 12 long; tarsus (107 – 112) about 1.9-2.1 longer than tibia; other length ratios, femur (70 – 77): genu (50 – 52): tibia (52 – 60), about 1.3: 0.9: 1.0. Legs II, III with tarsus/tibia length ratio about 1.8-2.0, leg IV with that ratio about 1.9-2.0. Chaetotactic formulae of leg segments as given for species-group. Tarsi II-IV with ventroapical process (7 – 9) bluntly pointed, smooth apically (Fig. 14D), and with setae (d-1) (15 – 20) nearly as long as pretarsus to base of claws (17 – 20). Tarsi II-III with paradactyli reaching tips of tarsal claws, but tarsus IV with anterior paradactylus extending about 3-5 beyond claw (Fig. 14D). Tarsi II (Fig. 13F), III with setae (l-1) shorter (16 – 19), more spinelike than (v-1) (28 – 34); dorsal seta md (34 – 37) no longer than pd-2, pl-3, mv (34 – 35); setae al-2, ad-2 (36 – 41), pl-2 (43 – 44), al-3, (pd-3) (28 – 32); all tarsal setae smooth. Tarsus IV with distal setae (v-1) symmetrical in size (26 – 31), slenderer than somewhat spinelike (l-1) (15 – 22).
Adult male. Relative leg thicknesses and lengths, excluding pretarsi, similar to those of female, I 305 – 310, II 255 – 260, III 242 – 250, IV 302 – 305. Legs with segment length ratios as in female. Tarsus I pretarsus 10 – 15 to base of claws, claws 5 – 9 long. Tarsi II-IV with dorsoapical setal processes (d-1), ventroapical process and pretarsal structures formed as in female, pretarsus lengths 12 – 17 excluding claws (10) (Fig. 13E). Apical setal processes (d-1) (11 – 13) shorter than pretarsi to bases of claws (14 – 17). Dimorphic leg setal structures not apparent.
Type material — All specimens collected at the La Selva Biological Station, Heredia Province, Costa Rica (10°26´N, 84°01´W, elevation 50-150 m): HOLOTYPE: adult female, 12 June 1997, ex Heliconia sp. leaves, successional plots, coll. ALAS. PARATYPES: 1 adult female, 1 adult male, with same data as holotype; 1 adult female, 12 June 1997, ex coexistent adults of Cephaloleia belti Baly and C. ornatrix Donckier from Heliconia leaves, coll. ALAS; 3 adult females, 15 April 1994, ex adult Chrysomelidae, no host plant data, coll. M. Paniagua; 2 adult females, 18 February 1994, ex adult Chrysomelidae, no host plant data, coll. ALAS; 3 adult females, 1 adult male, date not indicated, ex adult Cephaloleia belti on Calathea leaf, coll. not indicated.
Host identifications — As noted for the records of Lasioseius serripes, the identities of the associated beetles given above are in accord with current molecular studies, which indicate that records of Cephololeia belti and C. ornatrix on Heliconia pogognatha are in accord with field diet records. While C. belti is a host generalist, C. ornatrix is a host specialist of Heliconia pogonantha (Staines & Garcia-Robledo 2014, Garcia-Robledo et al. 2017).
Etymology — The specific name is based on the Latin noun,‘fuscina’, indicating a three-pronged fork or trident, and refers to the elongated three-pronged gnathotectum characteristic of this species.
Remarks — Limited collection data indicate a possible host association preference for Cephaloleia belti, a plant host generalist. If so, the mite/host size would correlate, as adults of L. ‘fuscina’ are the smallest of the described mites of this species-group, and among species of Cephaloleia, beetles of C. belti are relatively small-sized as well, although those of C. ornatrix are mid-sized. However, collection data uncertainties must be kept in mind, as two of the collection samples for this species also included many adults of Lasioseius serripes, one of them as well with a few adults of L. cassidini which also co-occurred with fuscina in two other samples (Table 1). Tentatively, subject to further studies, we conclude that, at La Selva, Lasioseius fuscina may be associated regularly with the host generalist Cephaloleia belti and to some extent with the host specialist C. ornatrix on Heliconia pogonantha.
ZOOBANK: F8B79273-9C72-4212-816C-8BC080C933CE ![]()









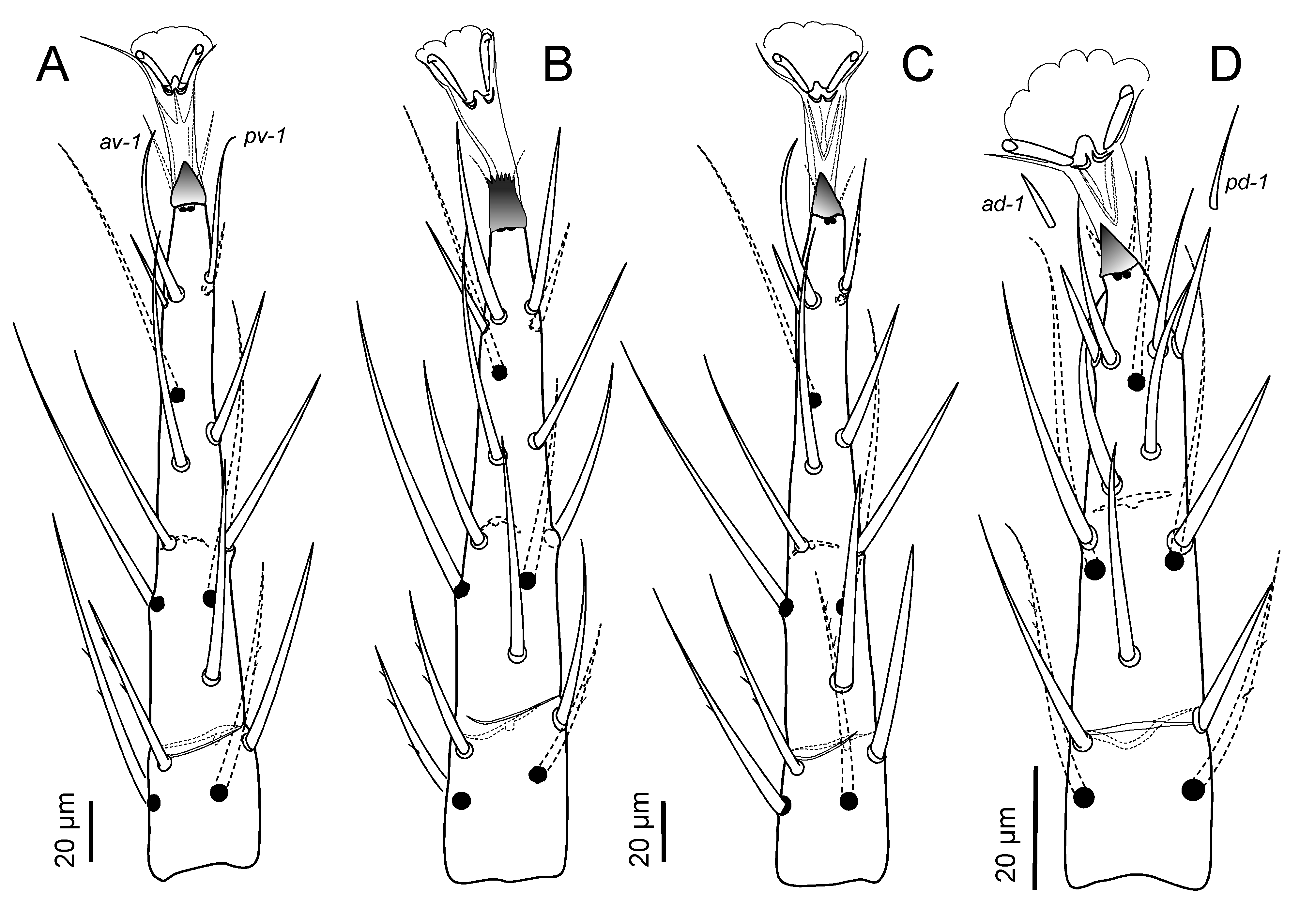
Diagnosis. Adult female: Sternal shield weakly lineate along lateral margins, smooth medially, its posterolateral corners not emarginated; poroids iv3 and setae st4 together on slender metasternal platelets; epigynal shield partially reticulated laterally, its lateral margins gently concave, narrowed anterior to setae st5; ventrianal shield with posterolateral margins on either side of anus nearly parallel, and with four pairs of opisthogastric setae, JV1 on shield; metapodal plates well separated on each side. Dorsal setae r6 on soft cuticle; setae Z5, S5 thickened, spinelike with blunt tips, Z4, S4 somewhat thickened but attenuated. Adult attributes: Gnathotectum anterior margin triramous, denticulate. Fixed chela with file of ca 20 teeth in female. Sternal setae st1 inserted on anterior edge of sternal shield. Tarsi II-IV with ventral apical process bluntly pointed, smooth apically; tarsi II-III with setae (v-1), (l-1) symmetrically stout; tarsus IV with apical seta av-1 longer, stouter than pv-1, (l-1), and pretarsus with anterior paradactylus longer than posterior one.
Idiosomatic dorsum — Adult female. Dorsal shield 540 – 595 long, 293 – 319 wide at level of r3, 320 – 345 at level of r5 (n=4), lightly reticulated over most of surface, but nearly smooth in dorsal hexagonal area (delimited by setae j5-z5-j6), not punctate behind level of setae Z4, but finely transversely striate between Z5 behind J5; shield with 37 pairs of setae, with lengths of most podonotal setae and J1-J3 not reaching bases of following setae; Z5, S5 thickened, spinelike with blunt tips, Z4, S4 somewhat thickened but attenuated (Figs. 10A, B): j1 (34 – 42), j2 (31 – 40), j3 (39 – 43), j4 (28 – 34), j5 (30 – 37), j6 (38 – 50), z1 (24 – 27), z2 (33 – 40), z3 (40 – 43), z4 (40 – 50), z5 (30 – 38), z6 (37 – 40), s1 (29 – 35), s2 (38 – 50), s3 (43 – 50), s4 (40 – 51), s5 (45 – 52), s6 (50 – 57), r2 (40 – 50), r3 erect (38 – 42), r4 (44 – 57), r5 (47 – 57), J1 (33 – 38), J2 (33 – 42), J3 (43 – 52), J4 (40 – 57), J5 minute and barbed (7 – 9), Z1 (53 – 62), Z2 (58 – 62), Z3 (62 – 70), Z4 (80 – 87), Z5 blunt (85 – 103), S1 (48 – 59), S2 (55 – 67), S3 (58 – 69), S4 (63 – 75), S5 blunt (75 – 90). Lateral soft cuticle with eight pairs of smooth setae, including r6 (45 – 60), R1 (35 – 52), R2-R6 shorter (28 – 42) and ZV5 posteriorly (40) (Fig. 10C).
Idiosomatic venter — Adult female. Tritosternum base 26 – 35 long, 15 – 16 wide at its base, with paired laciniae fused for about 10 – 17 of their total length (112 – 118). Presternal region with circumscribed pair of weakly granulated areas with jigsawed lineation, sometimes flanked anteriorly by a pair of narrow slightly sclerotized strips. Sternal shield 115 – 127 long, with slightly concave posterior margin, 98 – 109 wide, its surface nearly smooth medially, weakly lineate along lateral margins; sternal setae st1-st3 attenuate, of similar length (21 – 25) (Fig. 10C). Sternal poroids iv3 and setae st4 (20 – 22) inserted together on small metasternal platelets (21 – 26 long). Endopodal strips narrow between coxae III and IV. Epigynal shield with lightly reticulated lateral areas separated by smooth area medially, its narrowly rounded hyaline anterior margin reaching or slightly overlapping posterior edge of sternal shield, its lateral margins slightly concave, narrowed anterior to setae st5 (20 – 24), and its posterolateral corners slightly emarginated behind st5: shield 178 – 197 long, 82 – 102 at widest level at posterior angulate corners, 70 – 75 at level of st5, and 54 – 62 at narrowest width between legs IV. Opisthosomatic venter with two pairs of well separated metapodal platelets, primary pair oval, 30 – 40 long, 15 – 19 wide, secondary pair small, greatest dimension transverse, 18 – 25. Ventrianal shield inversely subtriangular with rounded anterior corners, with posterolateral margins on either side of anus nearly parallel; surface entirely sculptured, with primarily transverse lineation changing to reticulation behind setae JV2-ZV2, except anal region conspicuously punctate on either side of postanal seta; shield wider (216 – 235) than long, including cribrum (176 – 197), 113 – 132 wide at anal region; shield with four pairs of ventral setae and glandular pores gv3: JV1(19 – 22), JV2 (22 – 25), JV3 (23 – 27), ZV2 (23 – 29); with paranal setae (17 – 20) shorter than postanal seta (36 – 45). Soft cuticle with five pairs of simple opisthogastric setae, ZV1 (22 – 24) anterior to ventrianal shield, ZV3 (22 – 26), ZV4 (25 – 33), JV5 markedly thicker (53 – 57) flanking the shield, and ZV5 somewhat aligned behind R6; four pairs of poroids (including ivp). Peritrematal-exopodal shield lineate posteriorly behind leg IV; peritreme extending to level between insertions of setae s1 and z1 (Fig. 10C). Spermathecal apparatus with cup-like calyx 40 long including embolus and 22 wide at distal margin (Fig. 10 F).
Gnathosoma — Gnathotectum anterior margin triramous, with three short denticulate prongs, lateral margin slightly serrate (Fig. 10D). Cheliceral shaft, excluding basal section, 161 – 182 long, with slender digits; dorsal seta blunt (10); fixed digit with tiny, slightly offset subapical tooth and row of 20-28 very fine teeth (Fig. 10G), pilus dentilus minute (3) at level of 4th or 5th apical tooth; movable digit (70 – 80) tridentate. Subcapitulum rows of deutosternal denticles each with numerous (ca 20-30) fine denticles, 6th and sometimes 7th slightly widest and with ca 35 and 30 teeth, respectively; subcapitular setae hp1 (41 – 45) nearly as long as capitular setae pc (48 – 50), hp3 shorter (30 – 32), but longer than hp2 (13 – 17) (Fig. 10E). Palp length (160 – 165); apical margin of palptrochanter with a single spine anterolaterally; palptrochanter setae dissimilar in length, v1 42 – 45 and v2 27 – 36 (Fig. 10E).
Legs — Legs I (503 – 562) slightly longer than legs IV (488 – 546), and shorter than dorsal shield; other leg lengths (excluding pretarsi): II 390 – 445, III 380 – 442. Leg I pretarsus with biarticulated shaft (25 – 27 to base of claws), claws 15 long; tarsus (142 – 167) about 1.8 longer than tibia; other length ratios, femur (88 – 105): genu (73 – 82): tibia (80 – 95), about 1.1: 0.9: 1.0. Legs II-III with tarsus/tibia length ratio about 1.8-2.2, leg IV with that ratio about 1.8-1.9. Chaetotactic formulae of leg segments as given for species-group. Tarsi II-IV with ventroapical process (8 – 10) bluntly pointed, smooth apically (Figs 13I, 14C), and with setae (d-1) (13 – 18) nearly as long as pretarsus to base of claws (15 – 18). Tarsi II-III with paradactyli reaching tips of tarsal claws, but tarsus IV with anterior paradactylus about 1.5 longer (Fig. 14C). Tarsi II and III with setae (l-1) and (v-1) symmetrically stout, pl-1 (18) and pv-1 (18 – 25) slightly longer than al-1 (15) and av-1 (17 – 20), respectively; md (43 – 50) reaching bases of claws; setae (d-2) symmetrically attenuate (45 – 53). Tarsus IV with apical seta av-1 longer (28 – 45), stouter than pv-1 (16 – 23); setae (l-1) symmetrically similar in length (13 – 15), md (55 – 61) and (d-2) (60 – 72) attenuate (Fig. 14C).
ADULT MALE. Unknown.
Type material — All specimens collected at the La Selva Biological Station, Heredia Province, Costa Rica (10°26´N, 84°01´W, elevation 50-150 m): HOLOTYPE: adult female, (CRI 002, 739028), 8 July 1997, ex adult Chelobasis perplexa Baly, no host plant data, coll. M Paniagua. PARATYPES: 1 adult female, February 1994, associated with adult Chelobasis sp. on leaf of Heliconia pogonantha, coll. E.E. Lindquist & ALAS; 1 adult female, 15 April 1994, ex adult Chrysomelidae, no host plant data, coll. M. Paniagua; 1 adult female, 12 June 1997, ex adult Cephaloleia aequilata Uhmann (misidentification, see below) on leaf of Calathea, coll. ALAS; 1 adult female, 12 June 1997, ex adult Cephaloleia erichsonii Baly on leaf of Calathea, coll. ALAS.
Host identifications — With one exception, the identities of the associated beetles given above are in accord with current molecular studies of their field diet records, which indicate that one haplotype of Chelobasis perplexa sensu stricto is on Heliconia pogonantha. Cephalolaeia erichsonii is a host generalist. Cephaloleia aequilata does not occur at La Selva, and specimens identified as such from there are, in all cases checked by DNA barcodes, C. dilaticollis Baly, which is a host generalist, often on Heliconia, Calathea and other host plants (Carlos Garcia-Robledo, personal communications 2017; Garcia-Robledo et al. 2017).
Etymology — The specific name is a Latinized adjectival combination of ‘duo-obtusi-setis’, referring to two pairs of characteristically blunt-tipped setae Z5 and S5 on the dorsal shield.
Remarks — One female asymmetrically lacks one of setae s2. One female asymmetrically has seta JV4 inserted on the ventrianal shield; otherwise, this setal pair is symmetrically absent, characteristically in all species described of this group.
One female at hand, from under elytra of Cephaloleia erichsonii, Corcovado National Park, May 1997, coll. D. Johnson, has been tentatively identified as Lasioseius duobtusisetis, but is excluded from the type material. It differs primarily from other females by having lengths of most podonotal setae and J1-J3 longer, reaching beyond bases of following setae; yet setae Z5, S5 are thickened, spinelike with blunt tips, and Z4, S4 somewhat thickened but attenuated, which are distinctive of this species.


Two aspects of the sparse collection data are notable, with two records of association with beetles of the genus Chelobasis, and two other records with beetles of two species of Cephaloleia from leaves of a different genus of host plant, Calathea. Tentatively, subject to further studies, we conclude that, at La Selva, Lasioseius duobtusisetis is associated regularly with generalist forms of Chelobasis perplexa and Cephaloleia erichsonii on leaves of various species of Heliconia (and C. erichsonii also on various species of Calathea and Goeppertia), and also to some extent with another generalist form, Cephaloleia dilaticollis on a similar variety of host plant genera (Staines & Garcia-Robledo 2014, Garcia-Robledo et al. 2017).
Based on viewing a few adult females each with a fully formed larva in its opisthosoma, species of the cassidini-group are larviparous. Gravid females presented no indications of nymphiparity, such as occurs rarely among insect-parasitic forms of gamasine mites (Lindquist et al. 2016).
Although species of Lasioseius generally disperse as adult females phoretic on arthropodan associates, this was not determined for members of the cassidini-group, as the behavior of adults of both sexes running off and on the beetles may differ from a phoretic behavior of remaining on the beetles upon their departure for other plant host resources.


Our thanks to Carlos Garcia-Robledo (University of Connecticut, Storrs) and Charles Staines (Smithsonian Institution, Washington, D.C.) for providing current observations, images and references regarding the hispine beetle associates of these mites, to Derek M. Johnson (Virginia Commonwealth University, Richmond) for access to mite material associated with hispine beetles collected and identified by him in Costa Rica, and to John T. Longino (University of Utah, Salt Lake City) for access to relevant sections of the ALAS (Arthropods of La Selva) collection database. The technical expertise and field work of Danilo Brenes, Ronald Vargas, Maylin Paniagua and Nelci Oconitrillo in their former roles as parataxonomists in Project ALAS for our studies dealing with mites associated with rolled-leaf hispine beetles is greatly appreciated. Their work and field work by one of us (EEL) at La Selva, Costa Rica, was supported by National Science Foundation Grants BSR-9025024, DEB-9401069 and DEB-9706976. Helpful and interesting comments by the reviewers were appreciated.
Athias-Henriot C. 1969 — Les organes cuticulaires sensoriels et glandulaires des Gamasides. Poroïdotaxie et adénotaxie — Bull. Soc. zool. France, 94: 485-492.
Athias-Henriot C. 1971a — Un progres dans la connaissance de la composition métamérique des gamasides: leur sigillotaxie idiosomale (Arachnida) — Bull. Soc. zool. France, 96. 73-85.
Athias-Henriot C. 1971b — La divergence néotaxique des Gamasides (Arachnides) — Bull. Sc. Bourgogne, 28: 93-106.
Athias-Henriot C. 1975 — Nouvelles notes sur les Amblyseiini II. Le relevé organotaxique de la face dorsale adulte (Gamasides Protoadenique, Phytoseiidae) — Acarologia, 17: 20-29.
Britto E.P.J., Lindquist E.E., de Moraes G.J. 2011 — Redescription of Lasioseius floridensis Berlese, 1916 (Acari: Mesostigmata: Blattisociidae), with notes on closely related species — Zootaxa, 2905: 1-15.
Chant D.A. 1963 — The subfamily Blattisocinae Garman (=Aceosejinae Evans) (Acarina: Blattsocidae Garman) (=Aceosejidae Baker and Wharton) in North America, with descriptions of new species — Can. J. Zool., 41: 243-305. doi:10.1139/z63-025 ![]()
Christian A., Karg W. 2006 — The predatory mite genus Lasioseius Berlese, 1916 (Acari,Gamasina) — Abh. Ber. Naturkundemus. Görlitz, 77: 99-250.
Evans G.O. 1963 — Observations on the chaetotaxy of the legs in the free-living Gamasina (Acari: Mesostigmata) — Bull. br. Mus. nat. Hist. Zool., 10: 275-303. doi:10.5962/bhl.part.20528 ![]()
Evans G.O. 1964 — Some observations on the chaetotaxy of the pedipalps in the Mesostigmata (Acari) — Ann. Mag. Nat. Hist. Series 13, 6: 513-527.
Evans G.O. 1969 — Observations on the ontogenetic development of the chaetotaxy of the tarsi of legs II–IV in the Mesostigmata (Acari) — In: Evans G.O. (Ed.) Proceedings of the 2nd International Congress of Acarology 1967. Akadémiai Kiadó, Budapest pp. 195-200.
Garcia-Robledo C., Erickson D.L., Staines C.L., Erwin T.L., Kress W.J. 2013 — Tropical plant-herbivore networks: reconstructing species interactions using DNA barcodes — PLoS ONE, 8: e52967. doi:10.1371/journal.pone.0052967 ![]()
Garcia-Robledo C., Horvitz C.C., Kress J.W., Carvajal-Acosta N., Erwin T.L., Staines C.L. (2017) — Experimental assemblage of novel plant-herbivore interactions: ecological host shifts after 40 million years of isolation — Biotropica, 49(6): 803-810. doi:10.1111/btp.12464 ![]()
Garcia-Robledo C., Horvitz C.C., Staines C.L. 2010 — Larval morphology and development, host plants, adult longevity, sexual dimorphism and notes on natural history in Cephaloleia rolled-leaf beetles (Coleoptera: Chrysomelidae: Hispinae) — Zootaxa, 2610: 50-68.
Garcia-Robledo C., Kuprewicz E.K., Staines C.L., Erwin T.L., Kress W.J. 2016 — Limited tolerance by insects to high temperatures across tropical elevational gradients and the implications of global warming for extinction. Proceedings of the Natonal Academy of Sciences, 113(3): 680-685.
Garcia-Robledo C., Staines C.L., Kress W.J. 2015 — A new species of bromeliad-feeding Cephaloleia Chevrolat (Coleoptera, Chrysomelidae, Cassidinae) from Costa Rica: evidence from DNA barcodes, larval and adult morphology and insect diets — ZooKeys, 477: 143-155.
Harrison L. 1915 — Mallophaga from Apteryx, and their significance; with a note on the genus Rallicola — Parasitology, 8: 88-100.
Johnson K.P., Bush S.E., Clayton D.H. 2005 — Correlated evolution of host and parasite body size: tests of Harrison's rule using birds and lice — Evolution, 59: 1744-1753.
Johnston D.E., Moraza M.L. 1991 — The idiosomal adenotaxy and poroidotaxy of Zerconidae (Mesostigmata: Zerconina) — In: Dusbábek F., Bukva V. (Eds.). Modern Acarology Vol. 2. Prague, Academia and The Hague, SPB Academic Publishing bv. pp. 349-356.
Karg W. 1993 — Raubmilben: Acari (Acarina), Milben Parasitiformes (Anachtinochaeta) Cohors Gamasina Leach — DieTierwelt Deutschlands, 59, 523 pp., Jena, Gustav Fischer.
Krantz G.W. 1962 — Acari, free-living Mesostigmata. II. The family Aceosejidae — Exploration du Parc National de la Garamba, Mission H. De Saeger, Fasc. 34, 30 pp.
Krantz G.W., Redmond B.L. 1987 — Identification of glandular and poroidal idiosomal systems in Macrocheles perglaber F. & P. (Acari: Macrochelidae) — Exp. Appl. Acarol., 3: 243-253.
Lindquist E.E. 1964 — A revision of mites of the subfamily Blattisocinae (Acarina: Blattisocidae) in America north of Mexico. Ph.D. dissertation, University of California, Berkeley — University Microfilms, Inc., Ann Arbor, Michigan, 64-2093. iii + 413 pp.
Lindquist E.E. 1971 — New species of Ascidae (Acarina: Mesostigmata) associated with forest insect pests — Can. Entomol., 103: 919-942. doi:10.4039/Ent103919-7 ![]()
Lindquist E.E. 1994 — Some observations on the chaetotaxy of the caudal body region of gamasine mites (Acari: Mesostigmata), with a modified notation for some ventrolateral body setae — Acarologia, 35: 323-326.
Lindquist E.E. 2001 — Poising for a new century: diversification in acarology — In: Halliday R.B., Walter D.E., Proctor H.C., Norton R.A., Colloff M.J. (Eds), Acarology: Proceedings of the 10th International Congress. CSIRO Publishing, Melbourne, pp. 17-34.
Lindquist E.E. 2003 — Observations on mites of the subfamily Platyseiinae, with descriptions of two new species of Platyseius from North America (Acari: Mesostigmata: Ascidae) — In: Smith I.M, (Ed), An acarological tribute to David R. Cook (From Yankee Springs to Wheeny Creek). Indira Publishing House, West Bloomfield, Michigan, pp. 155-182.
Lindquist E.E., Béthoux O., Robin N., Sidorchuk E. 2016 — The first case of mite-on-mite hyperparasitism? — In: Ferragut Pérez F. (President), 8th Symposium of the European Association of Acarologists, Universitat Politècnica de València, July 11th-15th 2016, p. 88.
Lindquist E.E., Evans G.O. 1965 — Taxonomic concepts in the Ascidae, with a modified setal nomenclature for the idiosoma of the Gamasina (Acarina: Mesostigmata) — Mem. Entomol. Soc. Canada, 47: 1-64. doi:10.4039/entm9747fv ![]()
Lindquist E.E., Hunter P.E. 1965 — Some mites of the genus Proctolaelaps Berlese (Acarina: Blattisociidae) associated with forest insect pests — Can. Entomol., 97: 15-32. doi:10.4039/Ent9715-1 ![]()
Lindquist E.E., Moraza M.L. 2010 — Revised diagnosis of the family Blattisociidae (Acari: Mesostigmata: Phytoseioidea), with a key to its genera and description of a new fungus-inhabiting genus from Costa Rica — Zootaxa, 2479: 1-21.
Lindquist E.E., Moraza M.L. 2012 — A new genus of fungus-inhabiting mites of the family Blattisociidae (Acari Mesostigmata Phytoseioidea) from Costa Rica, with an updated key to genera of the subfamily Blattisociinae — Redia, 95: 9-19.
Lindquist E.E., Moraza M.L. 2014 — Mites coexistent with neotropical hispine beetles in unfurled leaves of Heliconia: a new genus and family of the Ascoidea (Acari: Mesostigmata: Gamasina) — J. Nat. Hist., 48: 1611-1651. doi:10.1080/00222933.2013.877995 ![]()
Lindquist E.E., Moraza M.L. 2016 — A new genus of mites of the subfamily Platyseiinae associated with Azteca ant galleries in Cecropia trees in Costa Rica (Acari: Mesostigmata: Blattisociidae) — Acarologia, 56: 293-319. doi:10.1051/acarologia/20162242 ![]()
Lindquist E.E., Walter D.E., Krantz G.W. 2009 — Order Mesostigmata — In: Krantz G.W., Walter D.E. (Eds.) A Manual of Acarology, Third Edition. Texas Tech University Press, Lubbock. pp 124-232.
Lindquist E.E., Wu K.W. 1991 — Review of mites of the genus Mucroseius (Acari: Mesostigmata: Ascidae) associated with sawyer beetles (Cerambycidae: Monochamus and Mecynippus) and pine wood nematodes [Aphelenchoididae: Bursaphelenchus xylophilus (Steiner and Buhrer) Nickle], with descriptions of six new species from Japan and North America, and notes on their previous misidentification — Can. Entomol., 123: 875-927. doi:10.4039/Ent123875-4 ![]()
Mineiro J.LC., Lindquist E.E., de Moraes G.J. 2009 — Edaphic ascid mites (Acari: Mesostigmata: Ascidae) from the state of Sao Paulo, Brazil, with description of five new species — Zootaxa, 2024: 1-32.
Moraes G.J. de, Abo-Schnaf R.I.A., Pérez-Madruga Y., Sánchez L., Karmakar K., Ho C-C. 2015 — The Lasioseius phytoseioides species group (Acari: Blattisociidae): new characterization, description of a new species, complementary notes on seven described species and a taxonomic key for the group — Zootaxa, 3980(1): 1-41.
Moraes G.J. de, Britto E.P.J., Mineiro J.L. de C., Halliday B. 2016 — Catalogue of the mite families Ascidae Voigts & Oudemans, Blattisociidae Garman and Melicharidae Hirschmann (Acaria: Mesostigmata) — Zootaxa, 4112(1): 1-299. doi:10.11646/zootaxa.4112.1.1 ![]()
Moraza M.L., Lindquist E.E. 2011 — A new genus of fungus-inhabiting blattisociid mites (Acari: Mesostigmata: Phytoseioidea) from Middle America, with a key to genera and subgenera of the subfamily Blattisociinae — Zootaxa, 2758:1-25.
Moraza M.L., Lindquist E.E. 2015 — Systematics and biology of mites associated with neotropical hispine beetles in unfurled leaves of Heliconia, with descriptions of two new genera of the family Melicharidae (Acari: Mesostigmata: Gamasina: Ascoides) — Zootaxa, 3931(3): 301-351. doi:10.11646/zootaxa.3931.3.1 ![]()
Moraza M.L., Lindquist E.E. 2016 — New species and biological data for the neotropical genus Hispiniphis (Acari: Mesostigmata: Melicharidae) associated with hispine beetles, genus Cephaloleia, in unfurled leaves of Heliconia — Syst. & Appl. Acarol., 21: 85-118.
Schmitt M., Frank M. 2013 — Notes on the ecology of rolled-leaf hispines (Chrysomelidae, Cassidinae) at La Gamba (Costa Rica) — ZooKeys, 332: 55-69. doi:10.3897/zookeys.332.5215 ![]()
Sellnick M. 1941 — Einige Milbenarten C. L. Kochs — Zool. Anz., 133, 146-155.
Staines C.L. 2009 — A review of the genus Chelobasis Gray, 1832 (Coleoptera: Chrysomelidae: Cassidinae) — Zootaxa, 2033: 58-68.
Staines C.L. 2011 — Hispines (Chrysomelidae, Cassidinae) of La Selva Biological Station, Costa Rica — ZooKeys, 159: 45-65.
Staines C.L., Garcia-Robledo C. 2014 — The genus Cephaloleia Chevrolat, 1836 (Coleoptera: Chrysomelidae: Cassidinae) — ZooKeys, 436: 1-355. doi:10.3897/zookeys.436.5766 ![]()
Walter D.E., Lindquist E.E. 1989 — Life history and behavior of mites in the genus Lasioseius (Acari: Mesostigmata: Ascidae) from grassland soils in Colorado, with taxonomic notes and description of a new species — Can. J. Zool., 67: 2797-2813.
Walter D.E., Lindquist E.E. 1997 — Australian species of Lasioseius (Acari: Mesostigmata: Ascidae): the porulosus group and other species from rainforest canopies — Invertebr. Taxon., 11: 525-547. doi:10.1071/IT96003 ![]()



2017-05-15
Date accepted:
2017-08-04
Date published:
2018-01-25
Edited by:
Faraji, Farid

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Moraza, Maria L. and Lindquist, Evert E.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)