



Phytoseiid mites from Basilicata region (Southern Italy): species diversity, redescription of Typhloseiulus arzakanicus Arutunjan and a dichotomic key of the genus Typhloseiulus Chant and McMurtry 1994 (Parasitiformes, Phytoseiidae)
Tsolakis, Haralabos1 and Ragusa, Ernesto2
1✉ University of Palermo, Department of Agricultural and Forest Sciences, Laboratory of Applied Acarology Eliahu Swirski, Edifice 5A, Viale delle Scienze, 13 - 90128 Palermo, Italy.
2University of Palermo, Department of Agricultural and Forest Sciences, Laboratory of Applied Acarology Eliahu Swirski, Edifice 5A, Viale delle Scienze, 13 - 90128 Palermo, Italy.
2017 - Volume: 57 Issue: 4 pages: 805-821
https://doi.org/10.24349/acarologia/20174195ZooBank LSID: 59ADF2BB-4CE3-45E9-8167-74A409E57E88
Keywords
Abstract
Phytoseiid mites are the most important predators of phytophagous mites that are harmful to vascular plants in various natural or agricultural ecosystems. They are considered to be the first exploiters of the foliage habitat (McMurtry, 2010), and their importance as biocontrol agents has been fully demonstrated in the last decades (McMurtry, 1982; Pickett et al., 1987; Duso, 1992; Abad-Moyano et al., 2010; Schmidt et al., 2013). However, limited information is available on phytoseiid fauna associated with uncultivated plants both in agroecosystems and natural ecosystems. It is well known that wild plants give shelter and alternative foods for phytoseiid mites (Duso et al., 2010; Tsolakis et al., 2016) thus sustaining their populations and this is very important especially when wild plants are in the environs of cultivated ones because they can be a reservoir for these important biocontrol agents (Fauvel and Cotton, 1981; Duso et al., 1997, 2010; Tixier et al., 1998, 2000).
This work aims at giving new data on the composition and the distribution of phytoseiid species associated with both cultivated and uncultivated plants in the Basilicata region. This is the first report on the Phytoseiidae fauna in this region which is important from an agricultural point of view.
Surveys were carried out in different localities of the Basilicata region (Southern Italy) between 1976 and 2014, and collections were done using the branch-shaking method (Tsolakis and Ragusa, 1999). Phytoseiid mites, fallen on a black plastic table were preserved in alcohol 70%, cleared in Nesbitt solution and mounted on slides in Hoyer’s medium. A differential interference contrast microscope (DIC) Zeiss Axioplan was used for drawings and measurements. For the dorsal and ventral chaetotaxy the nomenclature used is that proposed by Lindquist and Evans (1965) as adapted by Rowell et al. (1978) for the family Phytoseiidae, while for the terminology of adenotaxy and the insemination apparatus we follow, with some changes, Wainstein (1973), Athias-Henriot (1975; 1977) and Beard (2001). The leg chaetotactic formulae are those proposed by Evans and Till (1979). All measurements are given in micrometers (µm). Specimens of all species are kept in the Acari collection of the Laboratory of Agricultural and Applied Acarology “Eliahu Swirski”, Department of Agricultural and Forest Sciences, University of Palermo (Italy).
The frequency of phytoseiid species collected in the Basilicata region, their host plants and localities of the surveys are reported in table 1, appendixes 1 and 2 and figure 1. Of the plants surveyed 61% were uncultivated (herbaceous 25%, shrubby 30.6% and arboreal 44.4%) and 39% were cultivated (herbaceous 8.7%, shrubby 4.3% and arboreal 87%) (Appendix 2). Phytoseiid mites showed a different distribution on the surveyed plants. Euseius finlandicus (Oudemans, 1915) was the most common species, equally distributed on spontaneous and cultivated plants. Other species, Typhlodromus (Typhlodromus) exhilaratus Ragusa, 1977, Kampimodromus aberrans (Oudemans, 1930), Typhlodromus (Anthoseius) cryptus Athias-Henriot 1960, Typhlodromus (Typhlodromus) pyri Scheuten, 1857, Typhlodromus (Anthoseius) rhenanoides Athias-Henriot, 1960 and Typhlodromus (Anthoseius) intercalaris Livshitz and Kuznetsov, 1972 were found mainly on wild plants with a range varying from 57.1 to 84.2%. On the contrary, Euseius stipulatus (Athias-Henriot, 1960), Phytoseius finitimus Ribaga, 1904 and Amblyseius andersoni (Chant, 1957) were associated mainly with cultivated plants. The remaining species were rarely collected on both wild and cultivated plants. Quercus spp. were the forest plants with the richest phytoseiid fauna (20 species), while among the cultivated ones the greatest number of phytoseiids was associated with Vitis vinifera (11 species), followed by Malus spp. (9 species) (Appendix 2).



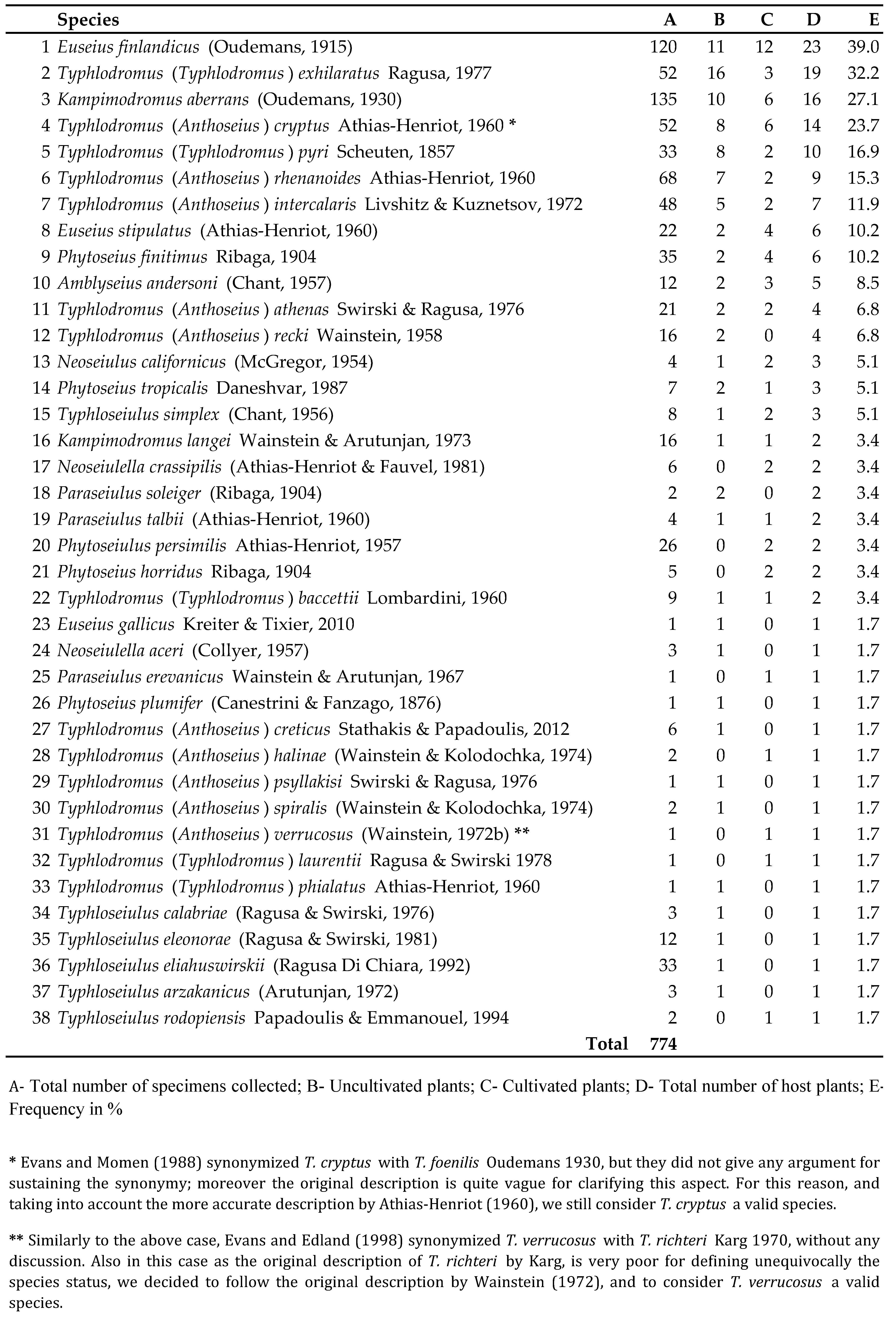
Phytoseiid species collected in the Basilicata region belong to eleven genera and two subgenera (Table 1). Among them, one of the most representative was the genus Typhloseiulus with 6 nominal taxa. We give here a new description of Typhloseiulus arzakanicus (Arutunjan), found for the first time in Italy, as the original one by Arutunjan did not report some important features currently used by taxonomists, and we also describe the male of this species.
(Figures 2 and 3)
Seiulus arzakanicus (original designation) — Arutunjan (1972)
Seiulus (Seiulus) arzakanicus — Beglyarov (1981)
Typhlodromus arzakanicus — Chant & Yoshida-Shaul (1983)
Typhloseiulus arzakanicus — Chant & McMurtry (1994)
Female (n =1)


Dorsum (Figure 2A) — Dorsal shield oval, strongly sclerotized and with evident reticulation throughout, without a distinct waist. Five pairs of minute solenostomes are visible on the dorsal shield: gd1 posteroantiaxial to j3, gd2 posteroparaxial to z4, gd6 anteroparaxial to Z1, gd8 anteroantiaxial to Z4 and gd9 anteroantiaxial to Z5. No poroids are visible on the dorsal shield, possibly due to the strong sclerotization of the shield. Some sigilla (muscle marks) are visible on podosoma (Figure 2). Dorsal setae of podosoma shorter than those of the opisthosoma. All setae are thick and inserted on tubercles. Setae j4, j5 and z5 are the shortest and subequal in length. Setae r3 and R1 on the interscutal membrane. All setae are smooth, except for Z5 which are lightly serrated. Measurements of dorsal and sublateral setae are as follows in µm; in parentheses measurements of the holotype (Arutunjan, 1972): j1 23 (24); j3 29 (24); j4 16 (15); j5 12 (12); j6 23 (21); J2 30 (25); J5 22 (15); z2 25 (18); z3 29 (30); z4 32 (32); z5 12 (12); s4 35 (36); s6 38 (42); Z1 35 (33); Z4 56 (72); Z5 60 (72); S2 42 (54); S4 61 (76); S5 10 (9); r3 17; R1 12. Angle Z1-gd6-j6 134°. Length of dorsal shield (j1-end of shield) 382, width at level of setae s4 203, width at level of setae S4 218.
Ventral idiosoma (Figure 2B) — Sternal shield sclerotized and smooth. Posterior margin slightly concave, anterior arms well visible. Setae st1, st2 inserted on the shield. Setae st3 and st4 tylochorous (on platelets); anterior platelets with outward projections. Poroids iv1 on the shield; poroids iv2 and iv3 on the platelets (Figure 2B). Length of sternal shield 46, width (st1-st1) 39, width (st2-st2) 50. Epigynial shield smooth; anterior margin of epigynium convex projecting laterally into two long arms. Lateral margin with a slight indentation posterior to st5. Genital sigilla (1-3 pairs) well visible. Posterior margin of the shield almost straight. Poroids iv4 on the interscutal membrane posteroantiaxial to st5. Genital sigilla of 4th and 5th pair well visible between bases of setae ZV1. Sigilla of 6th pair (sgpa) on the interscutal membrane, at level of setae JV1. Ventrianal shield (VAS) almost rectangular, sclerotized and irregularly ornamentated; only one pair of setae (JV2), besides circumanal setae are present on VAS. Solenostomes gv3 and muscle marks are not visible. Length of VAS 101 μm, width at level of JV2 59 μm, width at level of paranal setae 68 μm. Ratio length/width VAS at level of JV2 1.71. Ratio length/width at level of paranal setae 1.49. Six pairs of setae, ZV1, JV1, ZV2, ZV3, JV4 and JV5, on interscutal membrane. Setae JV5 smooth being the longest of the setae surrounding the VAS (21 μm). Anterior inguinal sigilla (metapodal platelets), 7 μm long; posterior inguinal sigilla 45 μm long and 6μm thick (Figure 2B).
Insemination apparatus (Figure 2F) — Receptaculum1 visible, slightly enlarged. Major duct cylindrical, narrow, lightly sclerotized, 57 µm long. Atrium bulbous, well differentiated, free and separate from the calyx by a constriction. Embolus2 well visible. Minor duct thin, clearly visible. Calyx campanulate, thick-walled all over, without basal neck, 12 µm long. Membranous vesicle well visible.
Legs (Figure 2C) — Seven setae are present on genu of leg II (1 1/1 2/0 2) and leg IV (0 2/1 2/1 1). One macroseta (18 – 20 μm) (sensu Beard 2001) is present on basitarsus of leg IV (17 in the holotype).
Chelicerae (Figure 2E) — Fixed digit with four teeth plus the apical tooth (three in the holotype); pilus dentilis evident. Movable digit with one tooth plus the apical one. 23μm long.
Peritreme (Figure 2D) — Apex of peritreme extends to bases of setae j1. Solenostome gd3 well visible on peritrematal shield at level of setae z4. Solenostome gdp crescentic, posterior to the opening of the stigma. The shape of peritreme presents the characteristic indentation found in other species of the genus (Figure 2D).
Male (Figure 3)


Dorsum (Figure 3A) — Dorsal shield oval, strongly reticulated as female. Five pairs of minute solenostomes are present on dorsal shield: gd1 posteroantiaxial to j3; gd2 anteroparaxial to z4; gd6 anteroparaxial to Z1; gd8 anteroantiaxial to Z4 and gd9 anteroparaxial to S5. Only poroids idm1, is1, idm3 and idm4 are visible on the dorsal shield. Dorsal setae thick, shorter than those of the female, smooth, inserted on tubercles. Setae of prosoma subequal in length than those of the opisthosoma. Setae r3 and R1 on dorsal shield. Peritrematal shield is fused with dorsal one at level of setae z3. Measurements of dorsal setae in μm means (min-max): j1 20 (19 – 20); j3 20 (20 – 21); j4 13 (12 – 14); j5 11; j6 14 (13 – 15); J2 18 (17 – 19); J5 14 (13 – 14); z2 17 (15 – 18); z3 20 (19 – 21); z4 21 (19 – 22); z5 9 (9 – 10); s4 22 (21 – 24); s6 20 (19 – 22); Z1 18 (17 – 19); Z4 28 (26 – 29); Z5 31 (29 – 33); S2 21 (20 – 22); S4 24 (22-25); S5 9 (8 – 10); r3 15 (14 – 15); R1 10 (9 – 11). Angle Z1-gd6-j6 53°. Length of dorsal shield (j1-end of shield) 290 (285 – 295), width at level of setae s4 174, width at level of setae S4 159 (154 – 164).
Ventral idiosoma (Figure 3B) — Sternogenital shield smooth. Cingulum3 absent. Ventrianal shield (VAS) almost completely ornamentated, triangular, length 108 μm (104 – 112), width at level of JV2 140 μm (137 – 142), width at level of paranal setae 76 μm (72 – 79). Setae JV5 11 μm long (10 – 11), smooth and tapered. Genital sigilla of 6th pair (sgpa) not visible. Solenostome gv3 not visible.
Legs — Seven setae are present on genu of leg II (1 1/1 2/0 2) and leg IV (0 2/1 2/1 1). One macroseta (15 – 17 μm) (sensu Beard 2001) is present on basitarsus of leg IV.
Chelicerae (Figure 3C) — Fixed digit with two teeth plus the apical one; pilus dentilis small but well visible. Movable digit with one bifid tooth in addition to apical tooth, 20 μm long (18 – 22). Spermatodactyl almost straight like a bird’s beak. Heel (antiramus) absent. Foot conical, finishing in a toe (processus apicalis) slightly rounded. Lateral projection not visible. A membranous velum slightly visible. Shaft (truncus) is thick; channel well visible along shaft and foot, 32 μm long (30 – 35).
Peritreme (Figure 3A) — Apex of peritreme between bases of setae j1 and j3. Peritrematal shield joint the dorsal one at level of setae z3. Solenostome gd3 on peritrematal shield.
Distribution — T. arzakanicus is reported from Armenia (Arutunjan, 1972), from Italy (present study) and from Serbia (Stojnić et al., 2014).
Remarks — The unique specimen collected on Quercus sp. is very close to T. arzakanicus. It differs from the latter species as some dorsal setae are shorter (Z4, Z5, S2 and S4) and some others are longer (j3, J2 and z2); moreover, five solenostomes are well visible on the dorsal shield, while they are not reported by Arutunjan (1972). The authors tried to get the type material of this species kept in the Scientific Center for Zoology and Hydroecology National Academy of Science of Republic of Armenia, but they never got the material. According to K.P. Dilbaryan, head of the above mentioned scientific center, the registration book of the Arutunjan’s collection is missing, and it is almost impossible to locate the slides of the type material, as well as the absence of written identification on the preserved slides (Dilbaryan pers. com).
Despite the above mentioned differences between our specimen and the holotype, we consider it as T. arzakanicus with waiting of more specimens and the possibility to observe the type material, in order to support the description of a new species.
Chant and Yoshida-Shaul (1983) created the simplex species group in the genus Typhlodromus. Some years later, Chant and McMurtry (1994) raised the simplex species group to the rank of genus, describing the new genus Typhloseiulus; at present 10 nominal species are known for this genus.
All Typhloseiulus species described up to now are of palearctic region: England, Portugal, Italy, Greece, Moldova, Armenia and Iran. The species of this genus have the same idiosomal setal pattern, 12-A:9B/JV-3:ZV and the same dorsal and ventral habitus: dorsal setae thick and thorn-like inserted on tubercles, often serrated and a strongly sclerotized and reticulated dorsal shield. Another characteristic of these species is the indented peritreme almost allover the marginal surface (Fig. 4a, b).
However, they differ by the shape of the calyx of the insemination apparatus. Accordingly, we decide to establish two new species groups:
A. eleonorae species group with the calyx of the insemination apparatus short, thick-walled and campanulate (Figure 4A, B): T. eleonorae and T. arzakanicus.

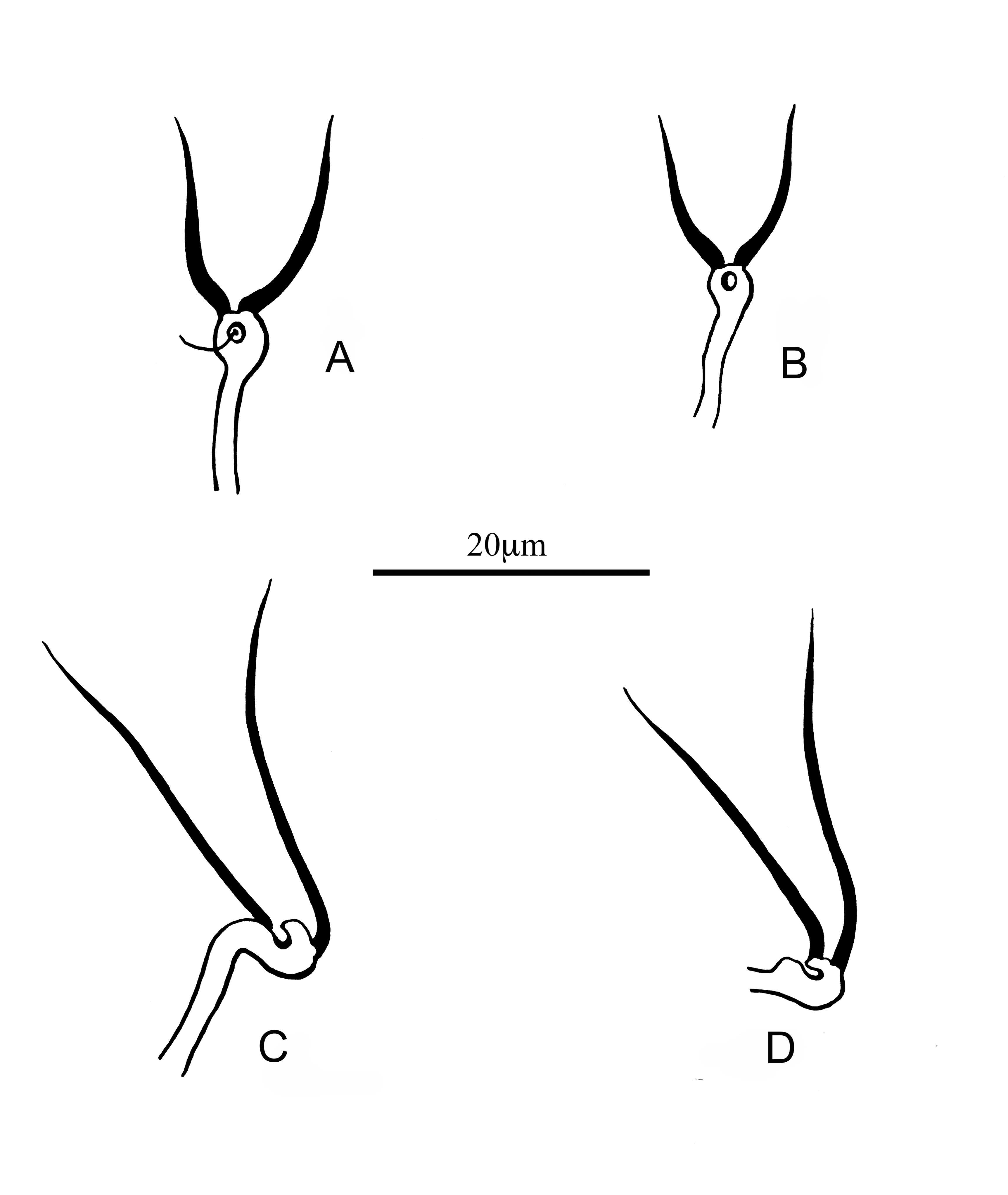
B. simplex species group with the calyx of the insemination apparatus elongate, flute shaped and flaring towards the distal part (Figure C, D): T. simplex, T. subsimplex, T. calabriae, T. peculiaris, T. carmonae, T. erymanthii, T. eliahuswirskii and T. rodopiensis.
1. Genu II with 7 setae. Setae j6 less than 30 µm and longer than setae S5 at least 2 times. All dorsal setae are smooth, except setae Z5 which are lightly serrated
...... T. arzakanicus
— Genu II with 8 setae. Setae j6 more than 40 µm and subequal to setae S5. Most of the dorsal setae are big, thick (6-8 µm in width) and strongly serrated
...... T. eleonorae
1. Genu II with 8 setae
...... 2
— Genu II with 9 setae
...... T. rodopiensis
2. Setae j6 short (less than 30 µm)
...... 3
— Setae j6 longer than 30 µm
...... 7
3. Setae J2 subequal to setae S5
...... T. calabriae
— Setae J2 longer than setae S5
...... 4
4. Setae j6 about half length of setae S5
...... T. subsimplex
— Setae j6 subequal or longer than setae S5
...... 5
5. Setae J2 four (or more) times longer than setae j6
...... T. peculiaris
— Setae J2 about double in length than setae j6
...... 6
6. Setae J2 three or more times longer than setae S5
...... T. simplex
— Setae J2 about two times longer than setae S5
...... T. erymanthii
7. Setae J2 about 2 times longer than setae j5
...... T. carmonae
— Setae J2 more than four times longer than setae j5
...... T. eliahuswirskii
Notes
1 Receptaculum – the proximal segment of the major duct after the opening between the 3rd and the 4th pair of coxae and may be simple or differentiate; in the latter case it is defined receptaculum (Athias-Henriot, 1977).
2 Embolus – the dimple present in the atrium on which the minor duct is inserted (Athias-Henriot, 1977).
3 Cingulum – in some species the end of the male’s peritrematal shield is attached to the anterior part of ventrianal shield through a narrow bridge of sclerotized tegument; this strip is defined cingulum.
Authors are deeply indebted to Mrs Eleonora Chiavetta and Peter Bartlett for the revision of the English text, and to Dr. K.P. Dilbaryan for the news about the Arutunjan’s collection. The “Progetti interni di Ateneo ID 2012-ATE-0452” provided financial support for this work.
Abad-Moyano R., Pina T., Pérez-Panadés J., Carbonell E.A., Urbaneja A. 2010 — Efficacy of Neoseiulus californicus and Phytoseiulus persimilis in suppression of Tetranychus urticae in young clementine plants — Exp. Appl. Acarol., 50: 317-328 doi:10.1007/s10493-009-9318-3 ![]()
Arutunjan E.S. 1972 — New species of mites of the genus Seiulus Berlese, 1887 (Parasitiformes, Phytoseiidae) — Dokl. Akad. Nauk Arm. SSR, 54(4): 237-240 (in Russian)
Athias-Henriot C. 1975 — Nouvelles notes sur les Amblyseiini. II – Le relevé organotaxique de la face dorsale adulte (Gamasides Protoadeniques, Phytoseiidae) — Acarologia, 27: 20-29.
Athias-Henriot C. 1977 — Nouvelles notes sur les Amblyseiini III. Sur le genre Cydnodromus : Redéfinition, Composition (Parasitiformes, Phytoseiidae) — Entomophaga, 22(1): 61-73. doi:10.1007/BF02372991 ![]()
Beard J. 2001 — A review of Australian Neoseiulus Hughes and Typhlodromips De Leon (Acari: Phytoseiidae: Amblyseiinae) — Invert. Taxon., 15: 73-158. doi:10.1071/IT99017 ![]()
Beglyarov G.A. 1981 — Keys to the determination of phytoseiid mites of the USSR — Inf. Bull. Int. Org. Biol. Contr. Nox. Anim. Plants, East Palaearctic Section, Leningrad, Russia: 2-97. [in Russian]
Chant D.A., Yoshida-Shaul E. 1983 — A world review of the simplex species group in the genus Typhlodromus Scheuten (Acarina: Phytoseiidae) — Can. J. Zool., 61(5): 1142-1151. doi:10.1139/z83-151 ![]()
Chant D.A., McMurtry J.A. 1994 — A review of the subfamilies Phytoseiinae and Typhlodrominae (Acari: Phytoseiidae) — Int. J. Acarol., 20: 223-310. doi:10.1080/01647959408684022 ![]()
Duso C. 1992 — Biological control of tetranychid mites in peach orchards of Northen Italy: role of Amblyseius andersoni (Chant) and Amblyseius finlandicus (Oud.) (Acari: Phytoseiidae) — Acta Phytopathol. Entomol. Hungarica, 27(1-4): 211-217.
Duso C., Malagnini V., Paganelli A. 1997 — Indagini preliminari sui rapporti tra pollini e Acari Fitoseidi — Allionia, 35: 229-239.
Duso C., Kreiter S., Tixier M-S., Pozzebon A., Malagnini V. 2010 — Biological control of mites in European vineyards and the impact of natural vegetation — In: Sabelis M.W., Bruin J. (Eds). Trends in Acarology: Proceedings of the 12th International Congress, p. 399-407. doi:10.1007/978-90-481-9837-5_65 ![]()
Evans G.O., Till W.M. 1979 — Mesostigmatic mites of Britain and Ireland (Chelicerata: Acari-Parasitiformes). An introduction to their external morphology and classification — Trans. Zool. Soc. London, 35: 139-270. doi:10.1111/j.1096-3642.1979.tb00059.x ![]()
Evans G.O., Momen F. 1988 — The identity of Seiulus rhenanus Oudms. and Typhlodromus foenilis Oudms. (Acari: Phytoseiidae) — J. Nat. Hist., London, 22: 209-216.
Evans G.O., Edland T. 1998 — The genus Anthoseius De Leon (Acari: Mesostigmata) in Norway — Fauna Norvegica, Ser. B , Norway, 45: 41-62.
Fauvel G., Cotton D. 1981 — Evolution des populations de tiphlodromes, Amblyseius aberrans (Oudms.) essentiellement, (Acariens: Phytoseiides) dans une haie d'ormes et un verger de pommiers et observations sur leur transport par le vent — Six. Journ. Phyt. Phytopharm. Circum-mediterraneennes, Perpignan (France) 25-28 Mai 1981, 471-479.
Lindquist E., Evans G.W. 1965 — Taxonomic concepts in the Ascidae, with a modified setal nomenclature for the idiosoma of the Gamasina (Acarina: Mesostigmata) — Mem. Entomol. Soc. Canada 47, 1-64. doi:10.4039/entm9747fv ![]()
McMurtry J.A. 1982 — The use of phytoseiids for biological control: Progress and future prospects. — In: Hoy M. A. (Ed). Recent Advances in Knowledge of the Phytoseiidae- University of California, Publication 3284, p. 23-48.
McMurtry J.A. 2010 — Concepts of classification of the Phytoseiidae: Relevance to biological control of mites. — In: Sabelis M.W., Bruin J. (Eds). Trends in Acarology: Proceedings of the 12th International Congress, p. 393-397. doi:10.1007/978-90-481-9837-5_64 ![]()
Pickett C.H., Gilstrap F.E., Morrison R.K., Bouse L.F. 1987 — Release of Predatory Mites (Acari:Phytoseiidae) by Aircraft for the Biological Control of Spider Mites (Acari:Tetranychidae) Infesting Corn — J. Econ. Ent., 80: 906-910. doi:10.1093/jee/80.4.906 ![]()
Rowell H.J., Chant D.A., Hansell R.I.C. 1978 — The determination of setal homologies and setal patterns on the dorsal shield in the family Phytoseiidae (Acarina: Mesostigmata) — Can. Ent., 110: 859-876. doi:10.4039/Ent110859-8 ![]()
Schmidt R.A., Beers E.H., Unruh T.R., Horton D.R. 2013 — Releases of Insectary-Reared Galendromus occidentalis (Acari: Phytoseiidae) in Commercial Apple Orchards — J. Econ. Ent., 106(5): 1996-2005. doi:10.1603/EC13153 ![]()
Stojnić B., Mladenović K., Marić I., Marčić D. 2014 — Species complexes of predatory mites and spider mites (Acari: Phytoseiidae, Tetranychidae) on cultivated and wild apple trees in Serbia — Int. J. Acarol., 40(7): 485-492. doi:10.1080/01647954.2014.956671 ![]()
Tixier M-S., Kreiter S., Auger P., Weber M. 1998 — Colonization of Languedoc vineyards by phytoseiid mites (Acari: Phytoseiidae): influence of wind and crop environment — Exp. Appl. Acarol., 22: 523-542. doi:10.1023/A:1006085723427 ![]()
Tixier M-S., Kreiter S., Croft B.A., Auger P. 2000 — Colonisation of vineyards by phytoseiid mites: their dispersal patterns in plot and their fate — Exp. Appl. Acarol., 24: 191-211. doi:10.1023/A:1006332422638 ![]()
Tsolakis H., Ragusa S. 1999 — Overwintering of phytoseiid mites (Parasitiformes, Phytoseiidae) on hazelnut (Corylus avellanae L.) in Sicily (Italy) — In: Bruin, van der Geest, Sabelis (Eds). Ecology and Evolution of the Acari, Kluwer Academic Publishers, p. 625-635.
Tsolakis H., Principato D., Jordà Palomero R., Lombardo A. 2016 — Biological and life table parameters of Typhlodromus laurentii and Iphiseius degenerans (Acari, Phytoseiidae) fed on Panonychus citri and pollen of Oxalis pes-caprae under laboratory conditions — Exp. Appl. Acarol., 70(2), 205-218. doi:10.1007/s10493-016-0076-8 ![]()
Wainstein B.A. 1972 — New species and subgenus of the genus Anthoseius (Parasitiformes, Phytoseiidae) — Zool. Zh., Russia, 51: 1477-1482. [in Russian]
Wainstein B.A. 1973 — On the structure of some organs of Phytoseiidae (Parasitiformes) important for taxonomy — Zool. Zh., 52(12): 1871-1872.


















2017-01-23
Date accepted:
2017-03-07
Date published:
2017-07-04
Edited by:
Kreiter, Serge

This work is licensed under a Creative Commons Attribution 4.0 International License
2017 Tsolakis, Haralabos and Ragusa, Ernesto
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)