



Predatory phytoseiid mites associated with cassava in Kenya, identification key and molecular diagnosis (Acari: Phytoseiidae)
Mutisya, Daniel L.1 ; El-Banhawy, El Sayed M. 2 ; Vicente dos Santos, Victor3 ; Kariuki, Charles W. 4 ; Khamala, Cannute P.M.. 5 and Tixier, Marie-Stéphane6
1✉ KALRO-Katumani, P. O. Box 340-90100, Machakos, Kenya and University of Nairobi, P. O. Box 30197-00100, Nairobi, Kenya
2University of Nairobi, P. O. Box 30197-00100, Nairobi, Kenya
3Montpellier SupAgro, UMR CBGP, 755 Avenue du Campus Agropolis, CS 30016, 34988 Montferrier-sur-Lez cedex, France
4KALRO-Katumani, P. O. Box 340-90100, Machakos, Kenya
5University of Nairobi, P. O. Box 30197-00100, Nairobi, Kenya
6Montpellier SupAgro, UMR CBGP, 755 Avenue du Campus Agropolis, CS 30016, 34988 Montferrier-sur-Lez cedex, France
2017 - Volume: 57 Issue: 3 pages: 541-554
https://doi.org/10.24349/acarologia/20174172ZooBank LSID: 30D9F6BD-A542-448F-8C72-F3B772DB33CF
Keywords
Abstract
Predatory mites of the family Phytoseiidae are the major natural enemies of the cassava green mites (CGM): Mononychellus tanajoa Bondar and M. progressivus Doreste (Acari: Tetranychidae) in Africa (Yaninek et al. 1993; Mutisya et al. 2015). The wet humid geographical regions are usually the most suitable for phytoseiid mites in cassava fields (Shipp and van Houten 1997; Walzer et al. 2007), as long dry periods cause high mortality of most phytoseiid species (De Courcy et al. 2004; Kariuki et al. 2005; Zundel et al. 2007). Bakker et al. (1993) reported that saturation deficit tolerance spectrum of phytoseiid species is an important factor for mite survival and breeding during periods of long drought. The phytoseiid mite Typhlodromalus aripo De Leon was introduced to Africa in the 1990’s to control the CGM. Following its release, it quickly spread to most warm-humid regions of the continent (Hanna et al. 1998; Kariuki et al. 2002; Zannou et al. 2007a). Since the release of T. aripo in the mid-1990s in Kenya’s coastal and western regions, full report on phytoseiid fauna on cassava was lacking in comparison to Malawi and Mozambique (Zannou et al. 2007a). Typhlodromalus aripo prefers inhabiting cassava apex and is reported to co-exist well with other native species present on leaves of the plant with no displacement impact of either species (Gnanvossou et al. 2005; Zannou et al. 2007a; Molo et al. 2016). ep The objective of the present study was to explore phytoseiid diversity in different cassava production zones. Further, it offered an opportunity to carry out a molecular diagnosis study of two species common on cassava. Finally, an identification key to species found on cassava in Kenya based on works of Moraes et al. (2001, 2007), Zannou et al. (2006, 2007b) and Chant and McMurtry (2006) was provided.
A survey was conducted (2011-2013) in 166 cassava fields of three geographical zones: the hot-dry low midlands (LM), the cool-wet upper midlands (UM) and the warm-wet coastal lowlands (CL) of Kenya (Table 1, Figure 1). In each field, 15 cassava plants were sampled at random. The phytoseiid specimens were collected by beating the plants with a wooden stick (60 cm long) over a blue plastic board for approximately two minutes. This technique was chosen as it allows collecting mites from both apices and leaves. With the aid of a head loop lens (magnification 4 x). Mites were then identified using morphological identification keys according to El-Banhawy and Knapp (2011), Moraes et al. (2001, 2007), Chant and McMurtry (2005) using a phase contrast microscope (x 400).




Specimens from plant apex in the coastal Kenya at Mtwapa (assumed to belong to T. aripo) and specimens from the middle canopy leaves at Embu (upper eastern region) were collected in 100 % alcohol vials for DNA analyses. Genomic DNA was individually extracted from four mite specimens using a Qiagen DNeasy tissue kit (Qiagen, Hilden, Germany), according to the DNA extraction protocol « Purification of Total DNA from Animal Blood or Cells » (Spin-Column Protocol) adapted for extracting total DNA from mites (Kanouh et al. 2010). After extraction, specimens were retrieved and mounted on slides according to the method developed by Tixier et al. (2010). A nuclear ribosomal gene section including ITS1-5.8S-ITS2 (reported as ITSS) and three mitochondrial markers (COI, 12S rRNA, Cytochrome B mtDNA) were used. For the COI mtDNA, two fragments were amplified, one using the former’s primers and one using the primers proposed by Navajas et al. (1996, 2000) and used in Okassa et al. (2009) and Tixier et al. (2006). Primers and thermal cycling are those reported in Tixier et al. (2012) and in Okassa et al. (2009). PCR products were sequenced along both strands using Dynamic ET Terminator Cycle Sequencing kit, and purified using ExoSAP-IT (Amersham). Sequences were aligned and analysed with Geneious v3.5.4 (Drummond et al. 2007). The genetic distances were calculated using the Kimura 2-parameter model using Mega 6.0.6® (Tamura et al. 2013).
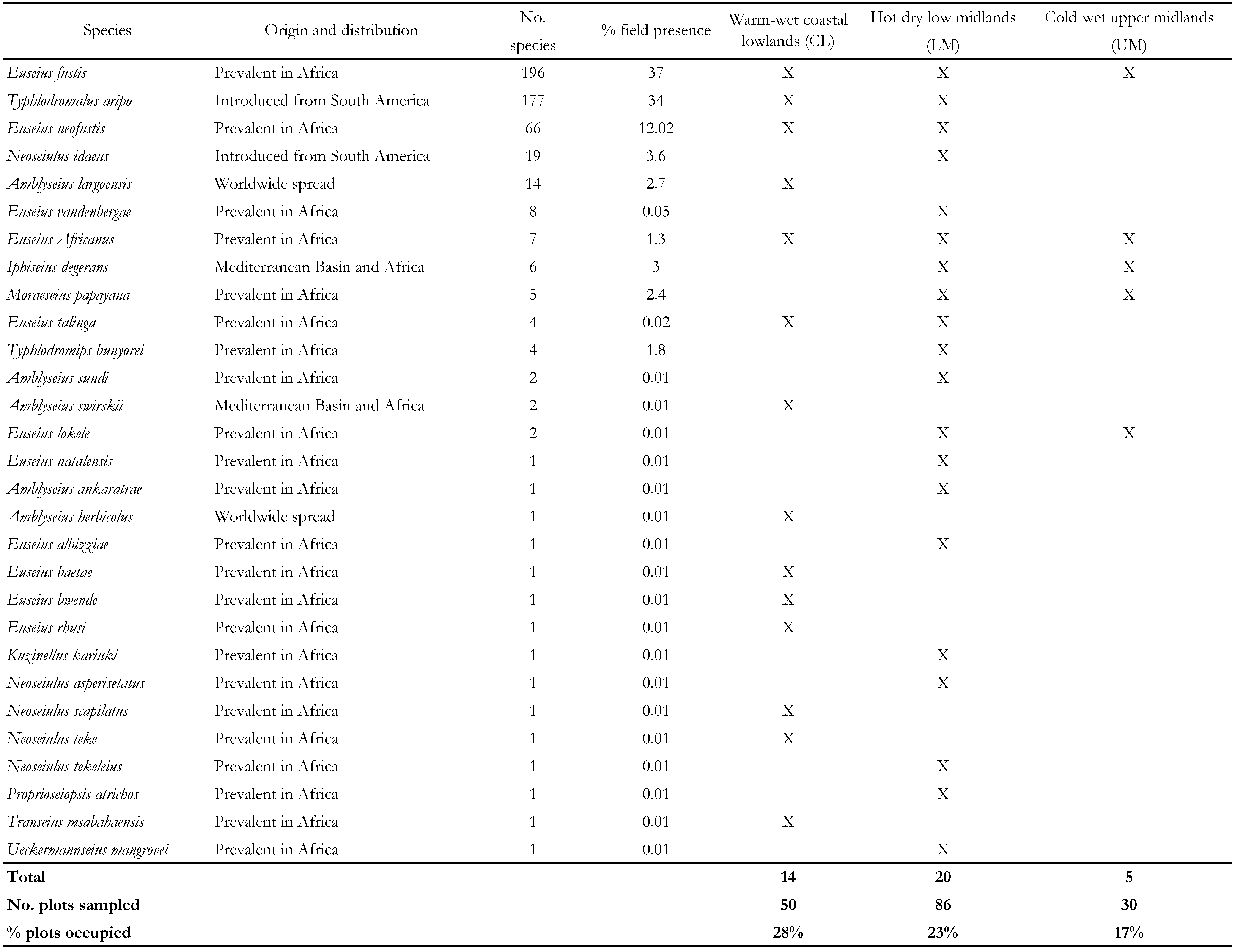
Within the 166 cassava plots sampled, 528 mites belonging to 29 species and 10 genera were identified (Table 2). The highest number of species belonged to genus Euseius Wainstein (12). The most frequent and numerous species were: E. fustis and T. aripo (present in 37 and 34% of sampled fields, respectively). All species found are reported below with some comments on DNA sequences when obtained. Information on the known distribution of the species is taken from the online ( http://www.lea.esalq.usp.br/phytoseiidae/ ![]() ) Phytoseiidae catalogue (Demite et al. 2014).
) Phytoseiidae catalogue (Demite et al. 2014).

space4.5mmAmblyseius ankaratrae Blommers, 1976
Specimens observed. 1 female: (01°21’50”S, 37°52’19”E, Alt. 1376 m Mwala Machakos, 31-V-2011).Previous records. Madagascar.Remarks. This is the first report of this species in the African continent.
Amblyseius herbicolus (Chant, 1959)
Specimens observed. 1 female: (04°08’56”S, 39°31’48”E, Alt. 25 m Kwale South Coast, 17-I-2012).Previous records. A cosmopolitan species.Remarks. This species is worldwide spread though mainly found in the Southern Hemisphere. This is the first record of this species on cassava.
Amblyseius largoensis (Muma, 1955)
Specimens observed. 14 females: (04°36’32”S, 39°27’7”E, Alt. 41 m Vanga South Coast, 17-I-2012; 04°45’40”S, 39°5’50”E, Alt. 25 m, 17-I-2011, Coast Kwale; 03°45’00”S 39°26’06”E, Alt. 10 m Malindi North Coast, 17-I-2011). Previous records. A cosmopolitan species. Remarks. Amblyseius largoensis was recovered in the coastal lowlands but at low frequency (0.4 %) with a total number of 14 specimens. This species is worldwide distributed and usually found in tropical climates. This is the first record of A. largoensis on cassava in Kenya. Earlier surveys have shown the presence of this species on mangrove trees in the same coastal region of Kenya (El-Banhawy and Knapp 2011).
Amblyseius sundi Pritchard & Baker, 1962
Specimens observed. 2 females: (03°42’36”S, 39°13’19” E Alt. 23 m Watamu North Coast, 17-I-2011).Previous records. Benin, Burundi, Cameroon, Cape Verde, Cuba, Democratic Republic of Congo, Ghana, Guadeloupe, Ivory Coast, Kenya, Madagascar Island, Malawi, Mozambique, Nigeria, Rwanda, Sierra Leone, Uganda, Zimbabwe.Remarks. This species has been mainly reported from the Ethiopian area and two areas in the Neotropical zone (Guadeloupe and Cuba). It has been reported once on cassava in Democratic Republic of Congo (Gutiérrez and Bonato 1994).
Amblyseius swirskii Athias-Henriot, 1962
Specimens observed. 2 females: (03°17’13”S, 39°2’38”E, Alt. 41 m Kilifi North Coast, 17-I-2011).Previous records. Argentina, Azerbaijan, Benin, Burundi, Cape Verde, Cuba, Democratic Republic of Congo, Egypt, Gaza Strip, Georgia, Ghana, Israel, Italy, Kenya, Malawi, Nigeria, Reunion Island (Kreiter et al. 2016), Saudi Arabia, Senegal, Spain, Tanzania, Turkey, USA, Yemen, Zimbabwe.Remarks. This species has been described from Israel and mainly recorded from Mediterranean Basin and Africa. It is a very efficient natural enemy commercialized and released in greenhouses for controlling phytophagous mites and thrips (Buitenhuis et al. 2015). It has been recorded once on cassava in Ghana (Zannou et al. 2007b).
Euseius africanus (Evans, 1954)
Specimens observed. 7 females: (04°43’30”S, 39°05’59”E, Alt. 32 m South Coast Lunga Lunga, 17-I-2011; 01°54’53” S, 37°42’26”E, Alt. 1116 m Kilala Machakos, 20-I-2011).Previous records. Burundi, Kenya, Malawi, Tanzania.Remarks. Euseius africanus was observed in 0.1% of the fields sampled in the regions of upper eastern, western and coastal Kenya. This species is only reported from the Ethiopian region. In Kenya, it has been already observed on other plants in coastal and central regions (El-Banhawy and Knapp 2011). This is the first report of this species on cassava.
Euseius albizziae (Swirski & Ragusa, 1978)
Specimens observed. 1 female: (00°37’33”S, 34°56’02”E, Alt.1266 m Ugunja Kisumu, 21-I-2011).Previous records. Kenya.Remarks. This species is only reported from Kenya and it is the first report on cassava.
Euseius baetae (Meyer & Rodriguez, 1966)
Specimens observed. 1 female: (04°43’37”S, 39°11’31”E, Alt. 26 m Kwale-Kokotoni South Coast, 17-I-2012). Previous records. Congo, Kenya, Malawi, Mozambique, South Africa.Remarks. This species is only recorded from Africa. Rodrigues (1968) reported it on cassava plants in Mozambique. Zannou et al. (2005) showed that this species was abundant on cassava plants in that country.
Euseius bwende (Pritchard & Baker, 1962)
Specimens observed. 1 female: (04°08’41”S, 39°27’57”E, Alt. 30 m Msabweni South Coast, 17-I-2012).Previous records. Democratic Republic of Congo, Kenya, Mozambique, Nigeria, South Africa, Zimbabwe.Remarks. This species is only reported from Africa. Zannou et al. (2005) reported this species as abundant in cassava plantations in Mozambique.
Euseius fustis (Pritchard & Baker, 1962)
Specimens observed. 196 females: (00°54’22”N, 34°26’60”E, Alt. 1689 m Kayenga Kakamega, 3-III-2013; 00°52’16”S, 37° 20’2”E, Embu, 19-II-2011; 00°25’01”S, 34°19’03”E, Alt. 1163 m Meru, 16-II-2011; 00°43’44”S, 37°23’46”E, Alt. 1407 m Rogo Migori 19-V-2011, 01°24’25”S, 37°29’46”E, Alt.1141 m Mukaa Machakos, 7-VII-2011; 03°10’48”S, 39°58’1”E, Alt. 43 m Mafueni North Coast, 18-I-2011; 04°36’32”S, 39°27’07” E, Alt. 41 m Vanga South Coast, 17-I-2012).Previous records. Benin, Burundi, Democratic Republic of Congo, Ghana, Kenya, Malawi, Nigeria, Rwanda, Tanzania, Uganda, Zimbabwe.Molecular sequences. Three to four sequences of this species were obtained for the DNA fragments 12S rRNA, COI (LCOHCO), CYB mtDNA and ITSS (Table 3). The genetic distances between the specimens of E. fustis are very low (12S rRNA: 0-0.002; CYTB mtDNA: 0-0.002; COI (LCOHCO): 0-0.032; ITSS: 0) and much lower than those previously reported from two species of a same genus (Tixier et al. 2006, 2008, 2011, 2012, 2014). Sequences of this species are not reported from Genbank.


Remarks. Euseius fustis was collected in 62% of the cassava fields sampled in Kenya. This species was mostly observed in eastern, western and coastal areas covering most agro-ecological zones of upper and low midlands (UMs & LMs) as well as the coastal lowlands (CLs). This species was already reported from different plants in western Kenya including cassava (Moraes and McMurtry 1988; Moraes et al. 1989; El-Banhawy and Kapp 2011). It is only present in Africa and has been reported on cassava also in Benin (Moraes et al. 1989), Malawi and Democratic Republic of Congo (Gutierrez and Bonato 1994; Zannou et al. 2005). The high densities presently observed as well as its high frequency indicate this is the most common phytoseiid on cassava in Kenya, even after the introduction of T. aripo as suggested by Onzo et al. (2014) due to partial niche separation among predator species (Onzo et al. 2010). Its ability to control M. tanajoa alone has been shown to be limited (Onzo et al. 2014; Mutisya et al. 2015).
Euseius lokele (Pritchard & Baker, 1962)
Specimens observed. 2 females: (00°42’18”S, 37°24’50”E, Alt. 1402 m Chuka Meru, 20-I-2011; 00°48’40”N, 34°42’36”E, Alt. 1219 mTeso Busia, 18-I-2012).Previous records. Benin, Burundi, Cameroon, Democratic Republic of Congo, Ghana, Kenya, Nigeria, Sierra Leone, Uganda.Remarks.This species is only recorded from Africa. It has been reported once on cassava in Democratic Republic of Congo.
Euseius natalensis (Van der Merwe, 1965)
Specimens observed. 2 females: (01°07’59”S, 037°27’50”E, Alt. 1141 m Kyanika Kitui, Date 27-I-2011).Previous records. Kenya, South Africa.Remarks. This species has only been recorded from Africa. It is its first report on cassava.
Euseius neofustis Moraes & McMurtry, 1988
Specimens observed. 66 females: (00°42’22”S, 37°15’50”E, Alt. 1401 m Egoji Meru, 19-V-2011; 00°27’47”S, 39°39’36”E, Alt. 28 m, Lunga Lunga, South Coast, 1-XII-2011; 03°17’06”S, 39°01’19”E, Alt. 23 m, Kilifi North Coast, 2-XII-2011).Previous records. Kenya, Tanzania, Thailand.Remarks. Euseius neofustis was observed in 20.2 % of the plots sampled. Sixty-six specimens were collected from eastern and coastal regions. El-Banhawy and Knapp (2011) reported the presence of this species on other plants in coastal region of Kenya and Moraes and McMurtry (1988) reported its occurrence on cassava in Kenya.
Euseius rhusi (Van der Merwe, 1965)
Specimens observed. 1 female: (03°02’10”S, 39°02’28”E, Alt. 29 m Kilifi North Coast, date 2-XII-2011).Previous records. Burundi, Colombia, Kenya, Mozambique, Reunion Islands, Rwanda, South Africa.Remarks. This species is mainly reported from Africa. Only one publication states its presence in Colombia. This is the first report of this species on cassava.
Euseius talinga (Pritchard & Baker, 1962)
Specimens observed. 4 females: (04°27’36”S, 39°14’44”E, Alt. 43 m Msabweni South Coast, 1-XII-2010; 00°52’16”S, 37°20’2”E, Alt. 1404m Embu, 19-II-2011).Previous records. Benin, Burundi, Democratic Republic of Congo, Ghana, Kenya, Malawi, Nigeria, Rwanda, Sierra Leone, Togo.Remarks. This species is prevalent in Africa, but it had never been reported on cassava before.
Euseius vandenbergae (Ueckermann & Loots, 1988)
Specimens observed. 8 females: (02°3’18”S, 37°19’11”E, Alt. 922 m Mwailu Makindu, 7-VII-2011)Previous records. Cameroon, Kenya, Rwanda, South Africa.Remarks. Euseius vandenbergae was rarely recovered in this study (8 specimens in 0.04 % of the fields sampled) in the coastal and western Kenya regions. It is the first time that E. vandenbergae is reported on cassava. This species was already observed in this country on other plants in coastal regions (El-Banhawy and Knapp 2011).
Iphiseius degenerans (Berlese, 1889)
Specimens observed. 6 females: (00°52’41”S, 37°20’06”E, Alt. 1402 m Embu, 20-V-2011; 02° 29’22”S, 037° 18’29”E, Alt. 1150m-Nzaui Makindu, 7-VII-2011).Previous records. Algeria, Benin, Brazil, Burundi, Cameroon, Canary Island, Cape Verde, Cyprus, Democratic Republic of Congo, Egypt, Georgia, Ghana, Greece, Israel, Italy, Kenya, Lebanon, Madagascar, Madeira Island, Malawi, Morocco, Nigeria, Portugal, Rwanda, Sierra Leone, South Africa, Sudan, Syria, Tanzania, Tunisia, Turkey, Uganda (Moraes et al. 2007), USA, Yemen, Zimbabwe.Remarks. This species is essentially reported from Africa, Middle East and southern Europe. It is used in greenhouses to control phytophagous mites and thrips (van Houten and van Stratum 1995; Vantornhout et al. 2004). This is the first report on cassava in Kenya; it has been already observed on this plant in Benin, Burundi, Malawi and Uganda (Zannou et al. 2005). Several studies report its low efficiency in controlling M. tanajoa because of the lack of spatial persistence in cassava fields (Nwilene and Nachman, 1996).
Moraeseius papayana (Van der Merwe, 1965)
Specimens observed. 5 females: (01°45’47”S, 37°05’20”E, Alt. 1316 m Machakos, 20-I-2011; 01°22’8”S, 37°30’43”E, Alt. 1587m-Nzaui Makindu, 00°42’16”S, 37°23’54”E, Alt. 1408 m Egoji Meru, 22-V-2011; 00°53’16”S, 37°18’02”E, Alt. 140 4m Siakago-Embu, 19-II-2011).Previous records. Kenya, Mozambique, South Africa.Remarks. Moraseius papayana was rarely observed (only 5 specimens in 0.1% of the fields sampled). This is the first report of this species in Kenya and on cassava.
Neoseiulus asperisetatus Zannou, Moraes & Oliveira, 2006
Specimens observed. 1 female: (01°12’25”S, 38°05’13”E, Alt. 1642 m Kyanika Kitui, 21-I-2011).Previous records. Benin, Cameroon, Democratic Republic of Congo, Kenya, Zambia.Remarks. his species is prevalent in Africa and has been previously reported from cassava in Benin (Zannou et al. 2006).
Neoseiulus idaeus Denmark & Muma, 1973
Specimens observed. 19 females: (01°20’51” S, 37°16’56” E, Alt. 1606 m Katumani Machakos 21-IV-2013; 02°45’43”S, 37°19’2”E, Alt. 922 m Kiboko Makindu 7-VII-2011; 01°13’24”S, 38°06’17”E, Alt. 1141 m Wikiliye Kitui, 21-I-2011; 00°10’55”S, 34°24’43”E, Alt. 1211 m Maseno Kisumu, 2-III-2011; 00°16’55”N, 37°00’50”E, Alt. 1402 m, Ishara Embu, 19-V-2011).Previous records. Argentina, Brazil, Colombia, Chile, Kenya, Paraguay, Peru, Venezuela.Molecular sequences. The COI sequences obtained matched with sequences of N. idaeus obtained by French researchers collected in Chile and Argentina (Table 3). DNA sequences of this species exist in Genbank but not for the fragment of COI herein sequenced.Remarks. This species was observed in 0.7 % of the cassava fields sampled, with a total of 19 specimens. Based on the morphological details given by El-Banhawy and Knapp (2011) for specimens from Kenya reported as Neoseiulus onzoi Zannou Moraes & Oliveira, we consider that those mites instead belong to N. idaeus. The latter species has been introduced to Africa between 1985 and 1993 to control M. tanajoa (Yaninek et al. 1992, 1993) without promising results (Yaninek et al. 1993; Kariuki et al. 2005). During 1988-1992, N. idaeus was released in coastal, western and eastern Kenya to control CGM, but recovery efforts did not yield any specimens (Yaninek and Herren, 1988; Yaninek et al. 1993). The specimens herein reported have been collected less than 500 meters from where N. idaeus was released in 1992 at KALRO Katumani field plot.
Neoseiulus scapilatus (Van der Merwe, 1965)
Specimens observed.1 female: (03°42’36”S, 39°13’19”E, Alt. 18 m Msabaha Malindi, 18-I-2011).Previous records. Benin, Burundi, Kenya, Malawi, Mozambique, Reunion Islands, Rwanda, South Africa, Uganda, Zimbabwe.Remarks. This species is widespread in Africa. It is the first record on cassava.
Neoseiulus teke (Pritchard & Baker 1962)
Specimens observed. 1 female: (03°17’06”S, 39°01’20”E, Alt. 29 m Kilifi North Coast, 2-II-2013).Previous records. Burundi, Democratic Republic of Congo, Ghana, Kenya, Malawi, Mozambique, Reunion Islands, Rwanda, Sierra Leone, South Africa, Tanzania, Zimbabwe.Remarks. This species is widespread in Africa. It was reported on cassava in Kenya (Moraes et al. 1989), Mozambique (Rodrigues 1968) and Malawi (Zannou et al. 2006). Nwilene and Nachman (1996) studied the reproduction characteristics of this species on M. tanajoa. It was more efficient than I. degenerans, but seems not efficient enough in field conditions (Nwilene and Nachman 1996).
Neoseiulus tekeleius El-Banhawy & Knapp, 2011
Specimens observed. 1 female: (04°21’04”S, 39°08’41”E, Alt. 36 m Ukunda South Coast, 17-I-2012).Previous records. Kenya.Remarks. This species has been only reported from Kenya on unknown plant.
Proprioseiopsis atrichos Moraes, Zannou & Oliveira, 2007
Specimens observed. 1 female: (01°10’41”S, 37°14’20”E, Alt. 1168 m Kilala Machakos, 20-II-2011).Previous records. Uganda.Remarks. It is the second record of this species after its description. This species is new for Kenyan fauna and has never been observed on cassava.
Transeius msabahaensis (Moraes & McMurtry, 1989)
Specimens observed. 1 female: (03°15’41”S, 39°32’10”E, Alt. 38m-Chumani Malindi, 16-XI-2011).Previous records. Kenya.Remarks. This species was described from Kenya and it is the third report after its description. It is the first report on cassava.
Typhlodromalus aripo De Leon, 1967
Specimens observed. 177 females: (04°27’46”S, 39°39’35”E, Alt. 44 m Lunga Lunga South Coast, 01°58’30”S, 37°45’57”E, Alt. 1584m- Kyambai Machakos, 12-II-2012; 03°11’22”S, 39°32’5”E, Alt. 34 m Gede Malindi North Coast, 12-IV-2013; 00°43’44”S, 37°23’46”E, Alt. 1407 m, Rogo Migori, 19-V-2011, 01°24’25”S, 37°29’46”E, Alt. 1141m, Mukaa Machakos, 7-VII-2011; 03°10’48”S, 39°58’01”E, Alt. 43 m, Mafueni North Coast, 2-II-2013; 02°45’43”S, 37°19’02”E, Alt. 922 m Kiboko Makindu 7-VII-2011).Previous records. Argentina, Benin, Brazil, Cameroon, Colombia, Democratic Republic of Congo, Costa Rica, El Salvador, Guadeloupe, Guyana, Jamaica, Malawi, Mexico, Mozambique, Paraguay, Peru, Trinidad, Uganda.Molecular sequences. One sequence of this species was obtained for the DNA fragments 12S rRNA and CYTB mtDNA (Table 3). The sequences of T. aripo match with those already in hand by French contributors of the paper were collected at Kiboko: 02°07’52”S, 37°25’57”E and Mtwapa: 03°10’01”S, 40°12’55”E , south east lowlands and coastal region of Kenya respectively.Remarks. Typhlodromalus aripo occurs in western, eastern and coastal regions and was absent in the cool upper eastern midlands (UM). Kariuki et al. (2002, 2005) has reported its continued occurrence on cassava since its release in Africa in 1990’s. Earlier reports have shown this species to be the most efficient in controlling CGM (Kariuki et al. 2005; Mutisya et al. 2014). This species was described from the Neotropical region and has been introduced to Africa for the control of the CGM where it suppresses pest populations to < 2 mites per leaf in most fields in optimum climatic conditions (Mutisya et al. 2015; Molo et al. 2016). In the absence of CGM, this species can survive on alternative preys such as whitefly and mealy bug nymphs (Gnanvossou et al. 2005; Mutisya et al. 2014).
Typhlodromips bunyorei El-Banhawy & Knapp, 2011
Specimens observed. 4 females: (00°07’19”S, 34°43’16”, Alt. 1411 m Mjini Kakamega, 3-III-2011).Previous records. Kenya.Remarks. Typhlodromips bunyorei was found in western Kenya. This is the first report on cassava. El-Banhawy and Knapp (2011) reported this species on other plants in western Kenya.
Ueckermannseius mangrovei El-Banhawy & Knapp, 2011
Specimens observed. 1 female: (01°43’58”S, 37°30’04”E, Alt. 155 8 m Tendelyani Machakos, 8-VII-2011).Previous records. Kenya.Remarks. This species is known from its original description from Kenya. This is the first report on cassava.
space4.5mmKuzinellus kariuki El-Banhawy & Knapp, 2011
Specimens observed. 1 female: (01°59’31”S, 37°33’32”E, Alt. 1339m-Mwala Machakos, 31-V-2011).Previous records. Kenya.Remarks. This is the second report of this species from Kenya, the first one being on coffee (El-Banhawy and Knapp 2011).
The most frequent and numerous species in this study were E. fustis (indigenous species) and T. aripo (introduced species), accounting respectively for 37 and 34 % of the specimens collected. They were present in more than 70% of the fields sampled. Euseius neofustis followed as a numerous species (13 % of the specimens collected). Each of the other species corresponded to less than 4 % of the specimens collected, most of them represented by a single specimen. Most of the species reported in this paper are indigenous to Africa and some of them are being reported first time on cassava in the present work. This study shows the spread of T. aripo and success of its introduction throughout Kenya.
Phytoseiid mites were observed from all climatic areas sampled. The highest number of plots occupied by Phytoseiidae was observed in the CL and LM zones corresponding to the most humid areas. The percentage of plots where phytoseiids were found were highest for the warm-wet low midlands and coastal lands; in the warm-drier coastal lowlands, 28 % of the plots were occupied by Phytoseiidae species. This observation validates the hypothesis that phytoseiids are susceptible to dry conditions. Only two species among the 29 sampled are observed in all regions (Euseius fustis and E. africanus) whereas 19 species were only found in one region, nine in the warm-wet coastal lowlands (CL) and ten in the hot-dry low midlands (LM). Only E. fustis was observed in the cool-wet upper midlands (UM). Typhlodromalus aripo prevailed in the CL (warm-wet coastal lowlands) region whereas E. fustis prevailed in the UM (cool-wet upper midlands) region.
ewcommandledetermination[2]oindent#1otfill#2
1. Podonotal region of the dorsal shield (anterior to setae R1) of the female with five or six pairs of “lateral” setae (j3, z2, z3, z4, s4, s6)
...... Kuzinellus kariuki
1’. Podonotal region of the dorsal shield (anterior to setae R1) of the female with four pairs of “lateral” setae (j3, z2, z4 and s4), z3 and s6 absent
...... 2
2 (1’). Sternal shield with median posterior projection, some forward “migration” of preanal setae ZV2 and/or JV2
...... 3
2’. Sternal shield without posterior projection, without forward “migration” of preanal setae ZV2 and/or JV2
...... 17
3 (2). Chelicera of normal size and shape, with prominent teeth evenly distributed along fixed digit; peritreme usually extending to level of seta j1
...... 4
3’. Chelicera reduced in size, stubby, with a group of small teeth clustered at the distal end of the fixed digit; peritreme usually not extending beyond seta j3, often much shorter
...... 5
4 (3). Dorsal shield setae short/minute (except Z5), shorter than distances between their bases; Z4 not as long as distance between its base and that of S4; dorsal shield smooth except for anterolateral striations.
...... Ueckermannseius mangrovei
4’. Dorsal shield setae of medium length, subequal; dorsal shield usually ornamented in addition to anterolateral striations; Z4 longer than distance between its base and that of S4
...... Typhlodromalus aripo
5(3’). Ventral and anal shields separated
...... Iphiseius degenerans
5’. Ventral, and anal shields fused
...... 6
6(5’). Seta Z1 present
...... 7
6’. Seta Z1 absent
...... Moraeseius papayana
7(6). Peritreme extending anteriorly to level of j1
...... 8
7’. Peritreme extending anteriorly at most slightly beyond level of j3
...... 10
8 (7). Calyx of spermatheca tubular
...... 9
8’. Calyx of spermatheca bifurcate, with a long neck
...... Euseius rhusi
9 (8). Calyx of spermatheca tubular but inflating progressively toward atrium
...... Euseius lokele
9'. Calyx tubular, with same diameter throughout
...... Euseius baetae
10 (7’). Peritreme extending anteriorly about to level of j3
...... 11
10’. Peritreme anteriorly reaching at most ca. half way between level of j3 and z2
...... 12
11(10). Some dorsal shield setae longer especially for setae j3 (40 – 50), S5 (24 – 27), Z1 (13 – 18), Z4 (18 – 21)
...... Euseius albizziae
11’. Some dorsal shield setae shorter, especially for setae j3 (27 – 38), S5 (8 – 13), Z1 (6 – 11), Z4 (8-11)
...... Euseius africanus
12(10’). Seta JV1 inserted on anterior margin of ventrianal shield
...... 13
12’. Seta JV1 inserted distinctly posterior to anterior margin of ventrianal shield
...... 14
13(12). Setae j3, z2 and z4 much less than half distances between their bases and bases of setae next behind
...... Euseius vandenbergae
13’. Setae j3, z2 and z4 ca. as long as distances between their bases and bases of setae next behind
...... Euseius natalensis
14(12’). Genu, tibia and tarsus of leg IV with several knobbed setae, dorsal shield reticulate
...... 15
14’. Genu, tibia and tarsus of leg IV at most each with one knobbed seta
...... 16
15(14’). Seta R1 rarely on dorsal shield, seta s4 ca. twice as long as distance between its base and pore immediately posterolaterad to it
...... Euseius fustis
15’. Seta R1 always on dorsal shield, seta s4 slightly longer than distance between its base and pore immediately posterolaterad to it
...... Euseius neofustis
16(14’). Seta j3 ca. half as long as seta j1, seta Z5 slightly longer than seta j1
...... Euseius talinga
16'. Seta j3 ca. as long as seta j1, seta Z5 slightly shorter than seta j1
...... Euseius bwende
17(2’). Seta J2 absent
...... Proprioseiopsis atrichos
17’. Seta J2 present
...... 18
18(17’). Macrosetae usually present only on leg IV (rarely missing on this leg) but sometimes also on leg III. Except for Z5, lateral dorsal setae subequal
...... 19
18’. Macrosetae usually present on legs II, III and IV, and sometimes also on leg I, lateral dorsal setae of quite different lengths
...... 24
19(18). GeIl without and GeIII rarely with a macroseta; fixed cheliceral digit usually with less than 6 teeth, rarely multidentate; never with these 2 character states together
...... 20
19’ GeIl and GeIII rarely without macrosetae; fixed cheliceral digit of usually with more than 6 teeth; most species with both of these 2 character
...... Typhlodromips bunyorei
20(19). Anterolateral setae (j3, z2 and z4) at least as long as distance between their bases and bases of respective immediate posterior setae
...... 21
20’. Anterolateral setae (j3, z2 and z4) shorter than distance between their bases and bases of respective immediate posterior setae
...... 23
21 (20). Dorsal setae, except j1 and J5 serrate; calyx of spermatheca abruptly constricted toward atrium
...... 22
21’. Dorsal setae, except Z4 and/or Z5 smooth; calyx of spermatheca not abruptly constricted toward atrium
...... Neoseiulus idaeus
22(21). Some dorsal setae shorter, especially j3=39, z4=48, Z5=58, seven teeth on fixed cheliceral digit
...... Neoseiulus teke
22’. Some dorsal setae longer, especially j3=55, z4 =70, Z5=75, four teeth on fixed cheliceral digit
...... Neoseiulus tekeleius
23 (20’). Seta Z1 at least 0.6x as long as distance between its base and base of S2; S2 at least 0.8x as long as distance between its base and base of S4
...... Neoseiulus scapilatus
23’. Seta Z1 at most 0.5x as long as distance between its base and base of S2; S2 at most 0.6x as long as distance between its base and base of S4
...... Neoseiulus asperisetatus
24 (18’). Ratios s4: Z1 and s4: S2 > 3.0
...... 25
24’. Ratio s4: Z1 > 3.0; ratio s4: S2 < 2.7
...... Transeius msabahaensis
25(24). Seta Z1 absent
...... Amblyseius sundi
25’. Seta Z1 present
...... 26
26(25’). Female ventrianal shield vase-shaped
...... 27
26’. Female ventrianal shield not vase-shaped (pentagonal)
...... Amblyseius swirskii
27(26). Ratio s4: Z1 ca. 2.0; Z4 shorter
...... Amblyseius ankaratrae
27’. Ratio s4: Z1 > 3.0; Z4 longer
...... 28
28(27’). Calyx of spermatheca trumpet-shaped
...... Amblyseius herbicolus
28’. Calyx of spermatheca tubular, parallel sided
...... Amblyseius largoensis
We acknowledge Ms Judith Shilavula, School of Biological Sciences, University of Nairobi, for proof reading the manuscript and the East Africa Agricultural productivity Project (EAAPP) for travel funds during the surveys. We also acknowledge the input of the anonymous reviewers to the final format of the paper.
Berlese A. 1889 — Acari, Myriopoda et Scorpiones hucusque in Italia reperta. Tipografia del Seminario — 6(54): 7-9.
Bakker F.M., Klein M. E., Mesa N.C., Braun A.R. 1993 — Saturation deficit tolerance spectra of phytophagous mites and their phytoseiid predators on cassava — Exp. Appl. Acarol., 17: 97-113.
Blommers L. 1976 — Some Phytoseiidae (Acarina: Mesostigmata) from Madagascar, with descriptions of eight new species and notes on their biology — Bijdragen tot Dierkunde, 46(1): 80-106.
Buitenhuis R., Murphy G., Shipp L. and Scott-Dupree C. 2015 — Amblyseius swirskii in greenhouse production systems: a floricultural perspective — Exp. Appl. Acarol. 65(4): 451-464. DOI: 10.1007/s10493-014-9869-9 doi:10.1007/s10493-014-9869-9 ![]()
Carrillo D., Hoy M.A., Pena J.E. 2014 — Effect of Amblyseius largoensis (Acari: Phytoseiidae) on Raoiella indica (Acari: Tenuipalpidae) by predator exclusion and predator release techniques — Florida Entomologist, 97(1): 256-26. doi:10.1653/024.097.0134 ![]()
Chant D.A. 1959 — Phytoseiid mites (Acarina: Phytoseiidae). Part I. Bionomics of seven species in southeastern England. Part II. A taxonomic review of the family Phytoseiidae, with descriptions of thirty-eight new species — Can. Entomol., Supplement 12: 1-166.
Chant D.A., McMurtry J.A. 2005 — A review of subfamily Amblyseiinae Muma (Acari: Phytoseiidae): Part VI. The tribe Euseiini tribe, subtribes Typhlodromalina n. subtribe, Euseiini n. subtribe and Ricoseiina n. subtribe — Int. J. Acarol., 31: 187-224. doi:10.1080/01647950508684424 ![]()
Chant D.A., McMurtry J.A. 2006 — A review of the subfamily Amblyseiinae Muma (Acari: Phytoseiidae): part IX. An overview — Internat. J. Acarol. 32(2): 125-152. doi:10.1080/01647950608684453 ![]()
De Courcy E., Kravar-Garde L., Fenlon J.S., Sunderland K.D. 2004 — Phytoseiid mites in protected crops: the effect of humidity and food availability on egg hatch and adult life span of Iphiseius degenerans, Neoseiulus cucumeris, N. californicus and Phytoseiulus persimilis (Acari: Phytoseiidae) — Exp. Appl. Acarol., 32: 1-13. doi:10.1023/B:APPA.0000018170.46836.11 ![]()
Demite P.R., McMurtry J.A., Moraes G.J. de 2014 — Phytoseiidae Database: a website for taxonomic and distributional information on phytoseiid mites (Acari) — Zootaxa, 3795: 571-577. doi:10.11646/zootaxa.3795.5.6 ![]()
Drummond A.J., Ashton B., Cheung M., Heled J., Kearse M., Moir R., Stones-Havas S., Thierer T., Wilson A. 2007 — Geneious v3.5 — Available from http://www.geneious.com/ ![]() . Accessed June 2016.
. Accessed June 2016.
El-Banhawy E.M., Knapp M. 2011 — Mites of the family Phytoseiidae Berlese from Kenya (Acari: Mesostigmata) — Zootaxa, 2945: 1-176.
Evans G.O. 1954 — The genus Iphiseius Berl. (Acarina: Laelaptidae) — Proc. Zool. Soc. London, 124: 517-526. doi:10.1111/j.1469-7998.1954.tb07793.x ![]()
Gnanvossou D., Hanna R., Yaninek J. S., Toko M. 2005 — Comparative life history traits of three neotropical phytoseiid mites maintained on plant-based diets — BioControl., 35: 32-39.
Gutiérrez J. 1987 — The cassava green mite in Africa: one or two species (Acari: Tetranychidae) — Exp. Appl. Acarol., 3: 163-168. doi:10.1007/BF01270477. doi:10.1007/BF01270477 ![]()
Gutierrez J., Bonato O. 1994 — Les acariens Tetranychidae attaquant le manioc au Congo et quelques-uns de leurs predateurs — J. Afric. Zool., 108(2): 191-200.
Hanna R., Yaninek J.S., Toko M., Onzo A., Gnanvossou D., Ojo D., Zannou L., Paraiso G. 1998 — Current status of cassava green mite Mononychellus tanajoa Bondar (Acari: Tetranychidae) biological control in Africa. In: the proceedings of the 7th Triennial Symposium of the International Society for tropical root crops — Africa Branch (ISTRC – AB)/ 11-17.
Kanouh M., Tixier,M.-S., Okassa M., Kreiter S. 2010 — Phylogenetic and biogeographic analysis of the genus Phytoseiulus (Acari: Phytoseiidae) — Zool. Scrip., 39: 450-461. doi:10.1111/j.1463-6409.2010.00439.x ![]()
Kariuki C. W, Toko M., Hanna R., Ngari B.M. 2002 — The progress in biological control of cassava green mite using phytoseiid predator Typhlodromalus aripo in Kenya — In: R. Hanna and M. Toko (eds) Proceedings of the regional meeting of the cassava green mite Biological Control Project, 15-17: 27-41.
Kariuki C. W., Hanna R., Toko M. 2005 — The impact of a predatory mite, Typhlodromalus aripo De Leon (Acari: Phytoseiidae) on cassava green mite population and yield of cassava in the field — Afr. Crop Sci. J., 7: 1401-1405.
Kreiter S., Vicente V., Tixier M.-S., Fontaine O. 2016 — An unexpected occurrence of Amblyseius swirskii (Athias-Henriot) in La Réunion Island (Acari: Phytoseiidae) — Acarologia, 56(2): 175-181. doi:10.1051/acarologia/20162254 ![]()
Meyer M.K.P., Rodrigues M.C. 1966 — Acari associated with cotton in southern Africa (with reference to other plants) — Garcia de Orta, Revista Junta Investigacoes de Ultramar, 13(2): 195-226.
Molo R., Aool, W., Adumo, S., Mutisya D.L. 2016 — Integrating cassava varieties and Typhlodromalus aripo to sustain biological control of cassava green mite — African Crop Science J., 24:117-126. doi.org/10.4314./acsj.v24i.13S
Moraes G.J. de, McMurtry J.A. 1988 — Some phytoseiid mites from Kenya, with description of three new species — Acarologia, 29(1): 13-18.
Moraes G.J. de, McMurtry J.A., Yaninek J.S. 1989 — Some phytoseiid mites (Acari, Phytoseiidae) from tropical Africa with description of a new species — Intern. J. Acarol., 15(2): 95-102. doi:10.1080/01647958908683830 ![]()
Moraes G.J. de, Zannou I.D., Ueckermann E.A., Oliveira A.R., Yanineck J.S., Hanna R. 2007 — Phytoseiid mites of the tribes Afroseiulini, Kampimodromini and Phytoseiulini, and complementary notes on mites of the tribes Euseiini and Neoseiulini (Acari: Phytoseiidae) from sub-Saharan Africa — Zootaxa, 1628: 1-22.
Moraes G.J. de, Ueckermann E.A., Oliveira A.R., Yaninek, J.S. 2001 — Phytoseiidae mites of the genus Euseius (Acari: Phytoseiidae) from Sub-Saharan Africa — Zootaxa, 3: 1-70.
Mutisya D.L., El-Banhawy E.M., Kariuki C.W., Khamala C.P.M. 2014 — Typhlodromalus aripo De Leon (Acari: Phytoseiidae) development and reproduction on major cassava pests at different temperatures and humidities: an indication of enhanced mite resilience — Acarologia, 54(4): 395-407.doi:10.105/acarologia/20142145
Mutisya D. L., El-Banhawy, E. M., C. P. M. Khamala, C. W. K. 2015 — Management of cassava green mite Mononychellus progresivus (Acari: Tetranychidae) in different agro-ecological zones of Kenya — Syst. Appl. Acarol., 20(1): 39-50. doi.org/10.11158/saa.20.1.5 doi:10.11158/saa.20.1.5 ![]()
Muma M.H. 1955 — Phytoseiidae (Acarina) associated with citrus in Florida — Ann. Entomol. Soc. Amer., 48: 262-272. doi:10.1093/aesa/48.4.262 ![]()
Navajas M., Guttierrez J., Lagnel, J. 1996 — Mitochondrial cytochrome oxidase I in tetranychid mites: a comparison between molecular phylogeny and changes of morphological and life history traits — Bull. Entomol. Res., 86: 407-417. doi:10.1017/S0007485300034994 ![]()
Navajas M., Fenton B. 2000 — The application of molecular markers in study of diversity in acarology: a review — Exp. Appl. Acarol., 24: 751-774. doi:10.1023/A:1006497906793 ![]()
Nwilene F.E., Nachman G. 1996 — Reproductive responses of Iphiseius degenerans and Neoseiulus teke (Acari: Phytoseiidae) to changes in the density of the cassava green mite, Mononychellus tanajoa (Acari: Tetranychidae) — Exp. Appl. Acarol., 20(5): 273-282. doi:10.1007/BF00052877 ![]()
Okassa M., Tixier M.-S., Cheval B., Kreiter S. 2009 — Morphological and molecular evidence for a new species status within the genus Euseius (Acari: Phytoseiidae) — Can. J. Zool. 87: 689-698. doi:10.1139/Z09-057 ![]()
Onzo A., Sabelis M.W., Hanna R. 2010 — Effects of ultraviolet radiation on predatory mites and the role of refuges in plant structures — Environ. Entomol. 39(2): 695-701. doi:10.1603/EN09206 ![]()
Onzo A., Sabelis M.W., Hanna R. 2014 — Single versus multiple enemies and the impact on biological control of spider mites in cassava fields in West-Africa — Exp. Appl. Acarol. 62(3): 293-311. doi:10.1007/s10493-013-9742-2 ![]()
Pritchard A.E., Baker E.W. 1962 — Mites of the family Phytoseiidae from Central Africa, with remarks on genera of the world — Hilgardia, 33: 205-309.
Rodrigues M.C. 1968 — Acarina de Mocambique: catalogo das especies relacionadas com a agricultura — Agronomia Mocambicana, 2(4): 215-256.
Swirski E., Ragusa S. 1978 — Three new species of phytoseiid mites from Kenya (Mesostigmata: Phytoseiidae) — Zool. J. Linn. Soc., 63: 397-409. doi:10.1111/j.1096-3642.1978.tb02101.x ![]()
Tamura K., Stecher G., Peterson D., Filipski A., Kumar S. 2013 — MEGA6: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods — Mol. Biol. Evol. 30: 2725-2729. doi:10.1093/molbev/mst197 ![]()
Tixier M.-S., Kreiter S., Barbar Z., Ragusa S., Cheval B. 2006 — The status of two cryptic species: Typhlodromus exhilaratus Ragusa and Typhlodromus phialatus Athias-Henriot (Acari: Phytoseiidae): consequences for taxonomy — Zool. Scrip., 35: 115-122. doi:10.1111/j.1463-6409.2006.00222.x ![]()
Tixier M.-S., Kreiter S., Croft B.A., Cheval B. 2008 — Kampimodromus aberrans (Acari: Phytoseiidae) from USA: morphological and molecular assessment of its identity — Bull. Entomol. Res., 98: 125-134. doi:10.1017/S0007485307005457 ![]()
Tixier M.-S., Okassa M., Liguori M.L., Poinso A., Salerno B., Kreiter S. 2010 — Voucher specimens for DNA sequences of Phytoseiid mites (Acari: Mesostigmata) — Acarologia, 50(4): 487-494. doi:10.1051/acarologia/20101984 ![]()
Tixier M.-S., Okassa M., Kreiter S. 2012 — An integrative morphological and molecular diagnostics for Typhlodromus pyri (Acari: Phytoseiidae) — Zool. Scrip., 41: 68-78. doi:10.1111/j.1463-6409.2011.00504.x ![]()
Tixier M.-S., Otto J., Kreiter S., Vicente V., Beard J. 2014 — Is Neoseiulus wearnei the Neoseiulus californicus of Australia? — Exp. Appl. Acarol., 62: 267-277. doi:10.1007/s10493-013-9740-4 ![]()
Tixier M.-S., Tsolakis H., Ragusa S., Poinso A., Ferrero M., Okassa M., Kreiter S. 2011 — An integrative taxonomical approach demonstrates the synonymy between Cydnodromus idaeus and C. picanus (Acari: Phytoseiidae) — Invert. System., 25: 273-281. doi:10.1071/IS11025 ![]()
Shipp J. L., van Houten Y.M., 1997 — Influence of temperature and vapour pressure deficit on survival of the predatory mite Amblyseius cucumeris (Acari: Phytoseiidae) — Environ. Entomol, 26: 106-113. doi:10.1093/ee/26.1.106 ![]()
Ueckermann E.A., Loots G.C. 1988 — The African species of the subgenera Anthoseius De Leon and Amblyseius Berlese (Acari: Phytoseiidae) — Entomology Memoir, Department of Agriculture and Water Supply, Republic of South Africa, 73: 1-168.
Van der Merwe G.G. 1965 — South African Phytoseiidae (Acarina). I. Nine new species of the genus Amblyseius Berlese — J. Entomol. Soc. South Africa, 28: 57-76.
Van Houten,Y.M., Van Stratum P. 1995 — Control of western flower thrips on sweet pepper in winter with Amblyseius cucumeris (Oudemans) and A. degenerans Berlese — In: Parker, B.L., Skinner, M. & Lewis, T. (Eds.), Thrips Biology and Management. NATO ASI Series A: Life Sciences Vol. 276, Plenum Press, New York: 245-248. doi:10.1007/978-1-4899-1409-5_37 ![]()
Vantornhout I., Minnaert H.L., Tirry L.,De Clercq P. 2004 — Effect of pollen, natural prey and factitious prey on the development of Iphiseius degenerans — BioControl, 49: 627-644.
Walzer A., Castagnoli M., Simoni S., Liguori M., Palevsk E., Schausberger P. 2007 — Intraspecific variation in humidity susceptibility of the predatory mite Neoseiulus californicus: survival, development and reproduction — Biol. Control., 41: 42-52. doi:10.1016/j.biocontrol.2006.11.012 ![]()
Yaninek J.S. 1994 — Cassava green mite intervention technologies — African Crop Science J., 2: 361-367.
Yaninek, J.S., Herren R.H. 1988 — Introduction and spread of the cassava green mite, Mononychellus tanajoa Bondar (Acari: Tetranychidae): An exotic pest in Africa and search for appropriate control methods — Bull. Entomol. Res. 79: 1-13. doi:10.1017/S0007485300016023 ![]()
Yaninek J.S., Onzo A., Ojo B. 1993 — Continent-wide experiences releasing neotropical phytoseiids against the exotic cassava green mite in Africa — Exp. Appl. Acarol., 16: 145-160.
Yaninek J.S., Mègevand B, de Moraes G. J., Bakker F., Braun A., Herren H.R. 1992 — Establishment of the neotropical predator Amblyseius idaeus (Acari: Phytoseiidae) in Benin, West Africa — BioControl Science and Technology, 1: 323-330. doi:10.1080/09583159109355211 ![]()
Zannou D.I., Hanna R., Moraes J.G, Kreiter S., Phiri G., Jone A. 2005 — Mites (Acari) of cassava (extslManihot esculenta
Crantz) habitat in Southern Africa — Int. J. Acarol. 31: 149-164. doi:10.1080/01647950508683667Zannou D.I., Moraes G.J. de, Ueckermann E.A., Oliveira A.R., Yaninek J.S., Hanna R. 2006 — Phytoseiid mites of the genus Neoseiulus (Acari: Phytoseiidae) from sub-Saharan Africa — Int. J. Acarol., 32(3): 241-276. doi:10.1080/01647950608684467 ![]()
Zannou I.D., Hanna R., Agboton B., de Moraes G.J., Kreiter S., Phiri G., Jones A. 2007a — Native phytoseiid mites as indicators of non-target effects of the introduction of Typhlodromalus aripo for the biological control of cassava green mite in Africa — Biol. Contr. 41: 190-198. doi:10.1016/j.biocontrol.2007.01.016 doi:10.1016/j.biocontrol.2007.01.016 ![]()
Zannou D.I., Moraes G.J. de, Ueckermann E.A., Oliveira A.R., Yaninek J.S., Hanna R. 2007b — Phytoseiid mites of the subtribe Amblyseiina (Acari: Phytoseiidae: Amblyseiini) from sub-Saharan Africa — Zootaxa, 1550: 1-47.
Zundel C., Hanna R., Scheidegger U., Nagel, P. 2007 — Living at the threshold: Where does the neotropical phytoseiid mite Typhlodromalus aripo survive in the dry season? — Exp. Appl.Acarol., 41: 11-26. doi:10.1007/s10493-007-9055-4 doi:10.1007/s10493-007-9055-4 ![]()



2016-07-28
Date accepted:
2016-12-06
Date published:
2017-05-17
Edited by:
Kreiter, Serge

This work is licensed under a Creative Commons Attribution 4.0 International License
2017 Mutisya, Daniel L.; El-Banhawy, El Sayed M. ; Vicente dos Santos, Victor; Kariuki, Charles W. ; Khamala, Cannute P.M.. and Tixier, Marie-Stéphane
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)