



Description of Halolaelaps coulsoni n. sp. (Acari, Halolaelapidae) from the High Arctic
Gwiazdowicz, Dariusz J.1 and Teodorowicz, Ewa2
1✉ University of Life Sciences, Department of Forest Pathology, Wojska Polskiego 71c, 60-625 Poznan, Poland.
2University of Life Sciences, Department of Forest Pathology, Wojska Polskiego 71c, 60-625 Poznan, Poland.
2017 - Volume: 57 Issue: 2 pages: 393-406
https://doi.org/10.1051/acarologia/20174164ZooBank LSID: A137B515-F6D2-4DCF-8AB7-1B325D60923B
Keywords
Abstract
Currently 80 species of the family Halolaelapidae mites have been described worldwide (Halliday 2008, Lindquist et al. 2009). They are common predators inhabiting a variety of microhabitats, but most commonly damp organic substrates including compost, manure and tidal debris. Several species of Halolaelaps have been found on marine algae and beach wracks in the intertidal zone. Halolaelapid deutonymphs of estuarine or intertidal habitats use amphipods for dispersal (Blaszak and Ehrnsberger 1993, 1995, 1998a, b, Karg 1993, Blaszak et al. 2001, Gwiazdowicz 2001).
Research was conducted in High Arctic on the island of Spitsbergen, Svalbard archipelago, which lies in the Norwegian Arctic between latitudes 74°N and 81°N and longitudes 10°E and 35°E, some 700 km north of mainland Norway. The archipelago has a land area of some 63,000 km2, of which 60% is under permanent ice and snow. The climate is relatively mild for the latitude due to the northern branch of the north Atlantic drift transporting considerable heat northwards. Nonetheless, the annual mean temperature is -6.7 °C with only four summer months, June to September, recording positive monthly averages, July being the warmest month at +5.5 ºC (Norwegian Meteorological Institute 2012). Monthly mean winter air temperature is often below -15 °C but daily minimum temperatures may decline to -40 °C on occasion. In the soil under snow, however, temperatures are often far less extreme and may be around -5 to -10 °C for the majority of the winter. In summer, the temperature of the upper soil layers may exceed air temperature considerably and reach over +20 ºC on warm days (Coulson et al. 1995).
Nowadays, 33 mesostigmatid mites have been recorded in Svalbard (Gwiazdowicz and Gulvik 2008, Ávila- Jiménez et al. 2011, Kolodochka and Gwiazdowicz 2014, Coulson et al. 2015). Two species of those belong to Halolaelapidae family, Halolaelaps gerlachi Hirschmann, 1966 and Saprosecans baloghi Karg, 1964 (Coulson et al. 2014).
During a study in the derelict mining town of Pyramiden in Svalbard an undescribed species of the Halolaelapidae family was collected. The aim of this paper is to describe the characteristics and ecology of the genus Halolaelaps as well as a detailed description of a species new to science.
Material were collected in Pyramiden on Spitsbergen from alongside a house wall where a nests of a black-legged kittiwake (Rissa tridactyla) were located. Mites were extracted from nest debris, litter and soil samples, in Tullgren funnels at the University Centre in Svalbard (UNIS) and preserved in 70% ethanol. Individuals were mounted in Hoyer's medium on glass slides and sealed with a nail polish for identification. All figures were drawn using a Zeiss Axioskop 2 microscope and Wacom UD-1212-R digitizer 2 tablet. Morphological details were measured as follows: setal length from base to tip, shield length along midline and width at the widest point of the shield. All measurements were taken in micrometers (μm). The chaetotaxy, symbols and the numbering system of setae on the dorsal and ventral side are after Evans (1963), Lindquist and Evans (1965) and Lindquist (1994).
Holotype and two paratypes of females and two paratypes of males and one deutonymph are deposited in the Poznan University of Life Sciences, Poland and one paratype of female and two males are deposited in Natural History Museum in London, United Kingdom.
Synonyms: Saintdidieria Oudemans, 1939; Saprolaelaps Leitner 1946
Type species: Gamasus marinus Brady, 1875 (= Halolaelaps glabriusculus Berlese et Trouessart, 1889)
The genus Halolaelaps was created by Berlese and Trouessart (1889), and later complimented by succeeding authors (Hirschmann 1966, Evans and Till 1979, Bregetova and Shcherbak 1977, Karg 1993, Halliday 2008). A typical character of Halolaelaps are two subequal dorsal shields, the idiosoma lacking horn-like caudal processes. The opisthonotal shield of many species bears a median incision running posteriorly from an anterior boarder towards the inner area of the shield. Dorsal setae commonly are simple or pilose. Tritosternum biflagellate with columnar base. Sternal shield subrectangular, fused with a presternal plate but not with endopodal plates, bearing three pairs of simple setae (st1-3) and two pairs of lyrifissures (iv1-2). Pair of st4 setae situated in a soft membrane or on the metasternal plates and pair st5 on a trapezoidal genital shield. Peritremes either short or long and peritrematal shields not expanded posteriorly; ventrianal shield broad, bearing 3-4 pairs of ventral setae or reduced to anal shield with three circumanal setae; metapodal plates varied in shape; palp genu with 6 setae, palp apotele 3-tined, corniculi horn-like. Chelicerae chelate-dentate; movable digit without excrescences. Tectum with a median prong either evenly trifurcate or denticulate.
Tarsus I with claws; trochanter I with 6 setae; femur I with 13 setae; genu I with 12 setae; tibia I with 11 setae; genu III with 9 setae; coxae sometimes with spines.
Remarks — A division of the genus Halolaelaps was made by Blaszak and Ehrnsberger (1993, 1995, 1998a,b) into 4 subgenera (Halolaelaps s.str., Halogamasellus, Haloseius, Saprogamasellus) based on the shape of epistome, the number of setae on femur I, and a spine on coxa II. Those designations and the characters used raised some multiple questions and controversies. For instance, why a type-species for Halolaelaps, that is Halolaelaps marinus, has not been placed into the subgenus Halolaelaps s.str. but into subgenus Hologamasellus (Blaszak and Ehrnsberger 1995) or why a type species have not been designated for the four subgenera that had been erected. All the controversial issues on this subject have been presented by Halliday (2008), and the authors of this paper agree with them completely. Therefore the discussion here does not follow the division of Halolaelaps into four subgenera.
Specimens examined — Holotype. Female. Spitsbergen, Pyramiden (78° 40'N, 016° 27'E), kittiwake nest detritus, July 16 2012, coll. S.J. Coulson and D.J. Gwiazdowicz; Paratypes: in the same location and date as holotype, two deutonymphs, coll. S.J. Coulson and D.J. Gwiazdowicz; three females, one deutonymph, four males in the same location, September 13 2012, coll. S.J. Coulson and A. Sjöblom.
Female (n=4), idiosoma oval, 690 – 730 μm in length and 430 – 450 μm in width.
ZOOBANK: A137B515-F6D2-4DCF-8AB7-1B325D60923B ![]()






Dorsal idiosoma — On dorsal side two shields of irregular edges - podonotal length 280 – 310 and width 340 – 380 μm and opisthonotal (280 – 310 x 300 – 340 μm) (Fig. 1A). Podonotal shield bearing 22 – 23 pairs of simple setae (6 per row "j", "z" and "s", and 4 or 5 setae in row "r"). Accordingly opisthonotal shield - usually with 14 pairs of simple setae. The total number of setae on both shields varies and this fact has been underlined in the section "Morphological variation". Lengths of setae on podonotal shield also vary, for example, that of "j" row are median in length (25 – 28 μm), while posterior setae in rows "z" and "s" (z5, z6, s5, s6) are noticeably longer (47 – 50 μm). The shortest are z1 (7 – 11 μm). Fine areolate ornamentation covers the shield in the area between "j" and "z" rows. Opisthonotal shield with a characteristic irregular incision in the middle (45 – 55 μm), starting from the anterior edge and running between setae J1-J1 and J2-J2. Similar to setae on podonotum, setae on opisthonotum differ in length e.g. J1-J3 are shorter (34 – 38 μm), slightly longer are J5, Z1, Z2 (41 – 45 μm), and the longest are Z4, S4 (57 – 58 μm). The anterior part of the shield between setae J1, Z1, S1 and J2, Z2, S2 is covered with delicate areolate ornamentation.
Ventral idiosoma — Tritosternum with elongate trapezoidal base (25 – 28 μm) and fine lightly pilose laciniae (42 – 47 μm) (Fig. 2A). A pair of lightly sclerotized pre-sternal plates fused with sternal shield. Sternal shield rectangular (105 – 115 x 80 – 90 μm) with three pairs of simple setae st1-st3 (21 – 25 μm) and two pairs of pores (Fig. 1B). Pair of setae st4 (29 – 32 μm) on soft integument. Epigynial shield peculiar (105 – 115 x 70 – 75 μm); epigynial setae on the edges of a shield, simple (30 – 32 μm); beneath epigynial (genital) shield, between setae JV1 a ZV1 lies a pair of narrow platelets (18 – 21μm). Three pairs of free narrow in shape endopodal plates present between coxae I/II, II/III and the largest between III/IV. Peritreme 260 – 280 μm long mounted on a broad peritrematal shield. Stigma on the horizontal line of coxae IV. Posteriorly to peritrematal shield lie two pairs of oval metapodal plates, a bigger outer platelets (21 – 25 x 10 – 12) μm and smaller inner platelets of 13 – 15 x 5 – 6 μm. Anal shield oval in shape (125 – 135 x 112 – 115 μm) with three typical circum-anal setae (45 – 48 μm). Between genital and anal shield are 8 – 9 pairs of simple setae (33 – 40 μm).
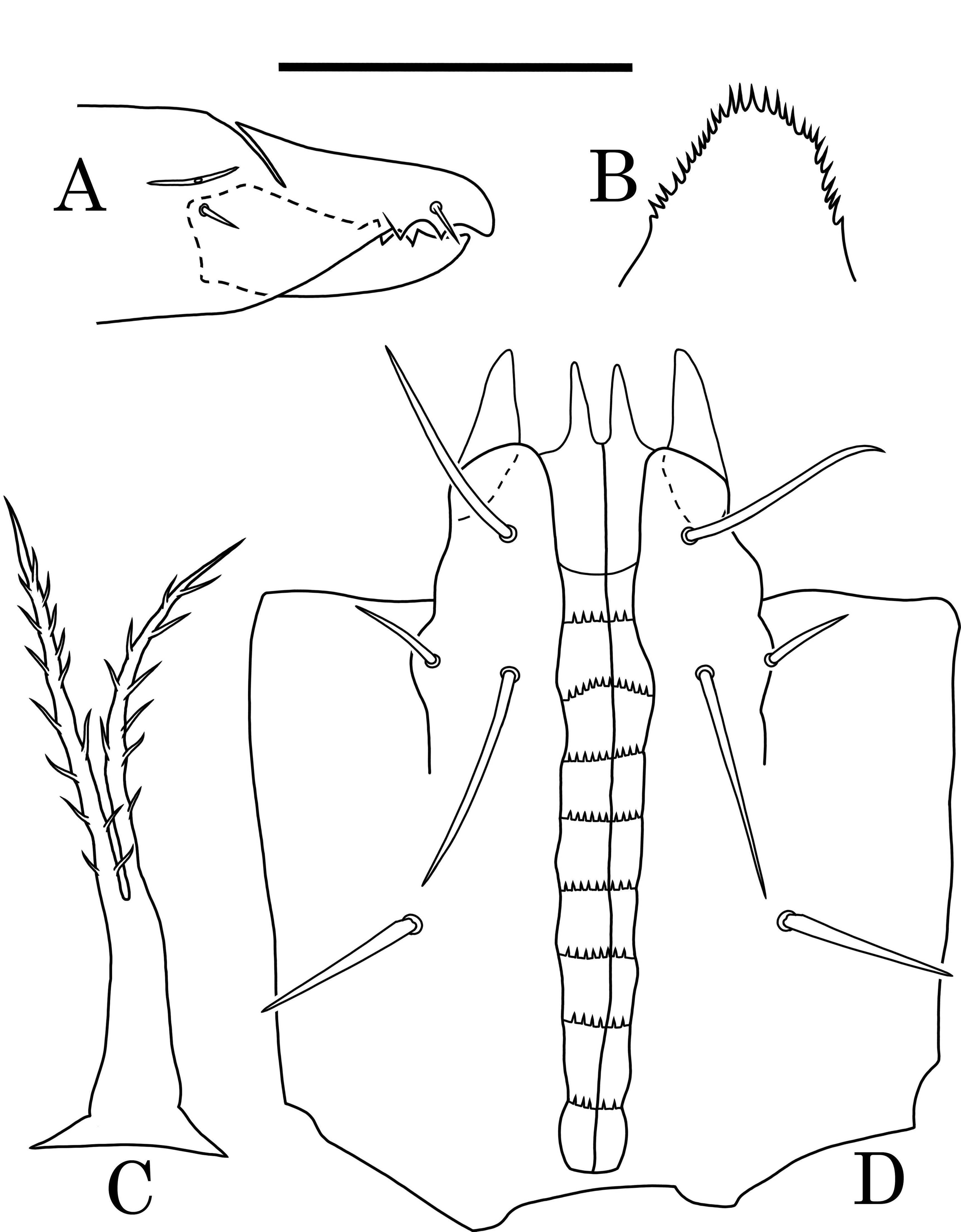
Gnathosoma — Corniculi short and robust, internal malae short, smooth. Palp coxal setae short (21 – 23 μm), internal posterior setae h3 longer (30 – 32 μm), external posterior setae h2 shorter (13 – 15 μm) and rostral setae h1 longer (28 – 31 μm). Hypostome with 8 transverse rows of denticles (8 – 15 denticles per row), denticles in each row irregular in size, and spacing (Fig. 2D). Tectum rounded with multiple small denticles (Fig. 2C). Chelicera with first and second segments slender, elongate (38 – 40 μm); fixed digit with two triangular distal teeth, pilus dentilis short, spine-like, dorsal cheliceral seta thick, prostrate. Movable digit slender (37 – 41 μm), with three distal triangular teeth (Fig. 2B). Palps 80 – 85 μm long (Fig. 2E, F, G), palp apotele three-tined.
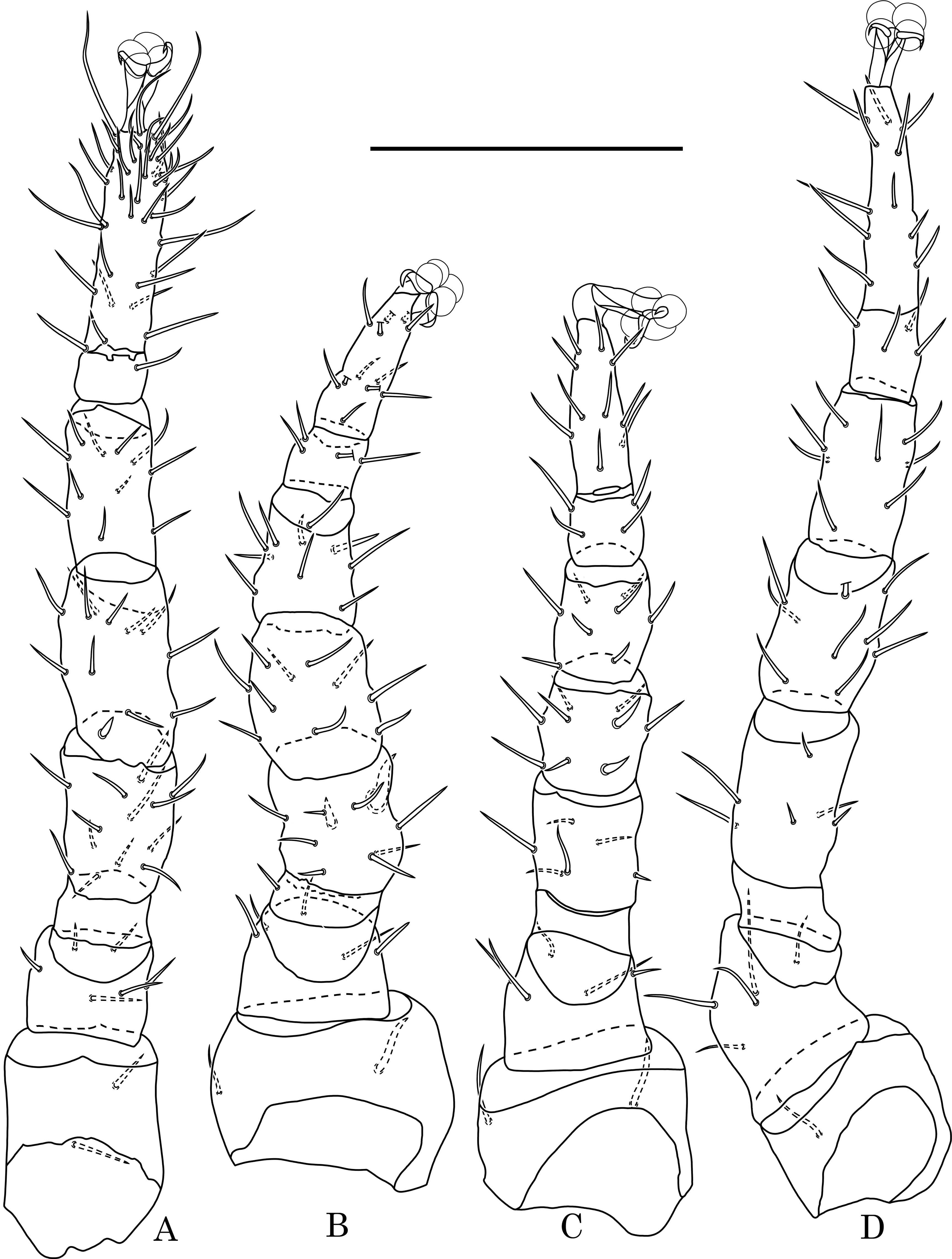
Legs — Variable in length, pretarsi equipped with claws and rounded pulvilli: I - 420 – 440 μm, II - 340 – 360 μm, III - 350 – 370 μm, IV - 430 – 450 μm. Chaetotaxy of legs is peculiar for genus Halolaelaps: leg I (coxa 2, trochanter 6, femur 13, genu 12, tibia 11), leg II (2, 5, 11, 10, 10), leg III (2, 5, 6, 9, 8), leg IV (1, 5, 6, 9, 8) (Fig. 3A, B, C, D).
Male (n=4), idiosoma oval, 470 – 520 μm in length and 310 – 340 μm in width.





Dorsal idiosoma — On a dorsal side two shields are present: podonotal (285 – 290 x 320 – 330 μm) with 23 pairs of simple setae and opisthonotal (200 – 210 x 290 – 300 μm) with 19 pairs of simple setae. Podonotal setae of varied lengths e.g. j1, j2, j6, z5 (28 – 32 μm), j3, j4, (37 – 38 μm) and s4, s5, s6 (44 – 46). Similar to the former, opisthonotal shield bears setae of different lengths e.g. J1, J2 (25 – 31 μm), J3 (21 – 24 μm), J4, J5 (17 – 21 μm), Z1, Z2, Z3, Z4, Z5 (36 – 42 μm), S1, S2, S3, S4 (45 – 51 μm). However, there is no anterior median incision in the opisthonotal shield which is present in the female. Both shields covered with delicate areolate ornamentation (Fig. 4A).
Ventral idiosoma — Tritosternum with elongate trapezoidal base (18 – 20 μm) and fine lightly pilose laciniae (31 – 34 μm) (Fig. 5A). Sternogenital shield long and narrow (220 x 120 – 125 μm) with five pairs of simple setae (23 – 26 μm) (Fig. 4B). Pre-sternal and endopodal plates fused with sternogenital shield. Genital opening at a height of coxae I/II. Peritreme 115 – 125 μm long and mounted on a broad peritrematal shield, stigma at a height of coxae IV. Pair of metapodal plates present, that of greater size (20 – 23 x 10 – 12 μm) and less size (12 – 14 x 6 – 8). Ventrianal shield with irregular edges bearing 5 – 6 pairs of simple setae (26 – 37 μm) and three even circum-anal setae (30 – 32 μm).
Gnathosoma — Corniculi short and robust, internal malae short and smooth. Palp coxal setae short (17 – 19 μm), internal posterior setae h3 longer (25 – 27 μm), external posterior setae h2 shorter (12 – 14 μm) and rostral setae h1 longer (26 – 28 μm). Hypostome with 8 transverse rows of denticles (6 – 9 denticles per row) all equal in size and spacing (Fig. 5D). Tectum rounded with multiple small denticles (Fig. 5C). Fixed digit of chelicera with two triangular distal teeth, pilus dentilis short, spine-like. Movable digit slender, with one distal triangular tooth and spermatodactyl (38 – 39 μm) (Fig. 5B). Palps similar to female's, slightly smaller (70 – 75 μm) (Fig. 5E, F, G).
Legs — Variable in length: I - 430 – 450 μm, II - 320 – 340 μm, III - 340 – 360 μm, IV - 440 – 450 μm. Chaetotaxy of male is similar to female: leg I (coxa 2, trochanter 6, femur 13, genu 12, tibia 11), leg II (2, 5, 11, 10, 10), leg III (2, 5, 6, 9, 8), leg IV (1, 5, 6, 9, 8) (Fig.6 A, B, C, D). The marked difference is the presence on femur II and genu III of two spiniform setae as well as that on coxae II and III there are not simple but singular spiniform setae.
Deutonymph (n=3), idiosoma oval, 480 – 520 μm in length and 320 – 340 μm in width.



Dorsal idiosoma — Dorsal side bearing two shields: podonotal (260 x 280 μm) with 23 pairs of simple setae and opisthonotal (220 x 260 μm) with 14 pairs of simple setae (Fig. 7A). Podonotal setae of varied lengths e.g. the shortest are z1 (9 μm), slightly longer j1 (20 – 22 μm), r2, r3 (25 – 27 μm), the remaining setae long (30 – 35 μm). The same variation can be seen on the opisthonotal shield where the shortest are J3, J4, J5 (14 – 17 μm), slightly longer than J1, J2 (21 – 24 μm), the remaining setae the longest (25 – 30 μm). As in the male, the opisthonotal shield lacks an anterior median incision. Both shields covered with delicate areolate ornamentation.
Ventral idiosoma — Tritosternum with elongate trapezoidal base (25 μm) and fine lightly pilose laciniae (45 μm) (Fig. 8C). Sternal shield long and narrow (220 x 120 – 125 μm) with five pairs of simple setae (26 – 31 μm). Pre-sternal plates combined with sternal shield. (Fig. 7B). Three pairs of narrow free endopodal plates present between coxae I/II, II/III, III/IV. Peritreme 225 – 230 μm long located on a wide peritrematal shield at height of coxae II/IV. Posterior of coxae IV lie oval metapodal plates (21 x 12 μm). Anal shield (88 – 91 μm x 95 μm) with three circumanal setae: two para-anals (20 – 21 μm) and one postanal (30 – 32 μm). Between the sternal and anal shields nine pairs of simple setae (21 – 26 μm) occur.
Gnathosoma — Corniculi short and robust, internal malae short, smooth. Hypostomatal setae of differentiated lengths: rostral setae h1 (23 – 24 μm), external posterior setae h2 (11 – 12 μm), internal posterior setae h3 (27 – 29 μm), palp coxal setae (20 – μm). Hypostome with 8 transverse rows of denticles (from 5 – 6 to 9 – 10 denticles per row) (Fig. 8D).
Tectum rounded with multiple small denticles (Fig. 8B). Chelicera the same as female (29 – 30 μm); fixed digit with two triangular distal teeth, pilus dentilis short. Movable digit slender, with three distal triangular teeth (Fig. 8A).
Legs — Variable in length: I - 380 – 400 μm, II - 310 – 330 μm, III - 310 – 220 μm, IV - 370 – 390 μm. Leg chaetotaxy in deutonymph is similar to female: leg I (coxa 2, trochanter 6, femur 13, genu 12, tibia 11), leg II (2, 5, 11, 10, 10), leg III (2, 5, 6, 9, 8), leg IV (1, 5, 6, 9, 8).
Halolaelaps coulsoni n. sp. is similar to Halolaelaps fallax (Hirschmann and Götz 1968) and Halolaelaps saproincisus (Hirschman and Götz 1968), due to the characteristic of opisthonotal shield: bears many pairs of setae, has an anterior incision in the middle as well as a similar type of tectum and, moreover, the female has an anal shield instead of ventrianal shield as in some species. However, Halolaelaps coulsoni n. sp. has certain features differentiating it from the species given above. H. saproincisus has some of the podonotal setae, and the majority of opistonotal setae, pilose and thus of a different type from those of both H. coulsoni n. sp. and H. fallax where all setae are simple (aciculate). At this point it should be underlined that setae under consideration go in different numbers - H. fallax bears 15 pairs of setae, but Halolaelaps coulsoni n. sp. only 14. Moreover, H. saproincisus on the lateral edges of opistonotal shield has incisions which are not present neither in Halolaelaps coulsoni n. sp. or H. fallax. Another differential character are the proportions of the anal shield: H. fallax has this shield wider than long, H. saproincisus has the shield of the same width and length, while in H. coulsoni n. sp. this shield is considerably longer than wide. H. cousoni n. sp. also has greater endopodal plates than the other Haloalelaps species herein. A final determining characteristics is the tectum: H. fallax and H. saproincisus have this triangular with teeth different in size totally reaching number of 10-15, while H. coulsoni n. sp. has all teeth similar in size and more numerous reaching about 25-30.
The morphological variability appears to concern mainly differences in the chaetotaxy. Four females have been analysed and none of them were free of inconsistencies, though different for each female. The most noticeable feature is the lack of setae in "J" row. One female lacks seta J2 on left side of shield and J1 on the right side (Fig. 1A); second and third females lack J2 on the right side, and the fourth female lacks both setae J2. First and second females have one setae Z5 laying outside opisthonotal shield but the third has this one on a shield however other seta (S4) present outside of it. On the podonotal shield some setae are absent or otherwise there are additional setae or the occurrence of some setae outside the shield. Morphological changes in chaetotaxy could be observed among leg setation as well. For example, usually on femur I there are 13 setae present but in one female there are 14 setae.
Despite this, morphological changes amongst males and deutonymphs represent not much variation. Some individuals lack of a single seta or have this displaced from the shield to the membrane. In addition, in one deutonymph on genu II instead of the typical 10 setae there are 11 setae.
A similar issue was noticed by Gwiazdowicz et al. (2011) after an analysis of 53 specimens (45 females and 8 males) of Arctoseius haarlovi Lindquist 1963, where 22 (18 females and 4 males, 41.5%), featured chaetotaxy variability. Although both species have been recorded in High Arctic, it is very unlikely that those changes could be due to a harsh environment conditions, as in other species of mesostigmatid mites from this geographical region such changes have not yet been observed, or at least not in a such numerous and frequent basis.
The species is dedicated to our friend, a scientist exploring the invertebrate fauna of the High Arctic, Prof. Dr. Stephen J. Coulson from University Centre in Svalbard, Longyearbyen, Norway.
The fieldwork was funded via Norwegian Research Council project AVIFauna (6172/ S30) and internal funding from the University Centre in Svalbard (UNIS).
Ávila-Jiménez M.L., Gwiazdowicz D.J., Coulson S.J. 2011 — The mesostigmatid mite (Acari: Parasitiformes) fauna of Svalbard: a revised inventory of a high Arctic archipelago — Zootaxa, 3091: 33-41.
Berlese A., Trouessart E. 1889 — Diagnoses d'Acariens nouveaux oupeuconnus — Bull. Bibl. Scient. Ouest, 2: 121-143.
Blaszak C., Ehrnsberger R. 1993 — Beiträge zur Kenntnis von Halolaelaps (Saprogamasellus) Götz,1952 (Acari: Gamasida: Halolaelapidae) — Genus, 4: 143-267.
Blaszak C., Ehrnsberger R. 1995 — Beiträge zur Kenntnis von Halolaelaps (Halogamasellus) subgen. nov., (Acari: Gamasida: Halolaelapidae) — Osnab. Nat. Mit., 20/21: 25-94.
Blaszak C., Ehrnsberger R. 1998a — Beiträge zur Kenntnis von Halolaelaps (Halolaelaps s. str.), (Acari: Gamasida: Halolaelapidae) — Osnab. Nat. Mit., 24: 159-181.
Blaszak C., Ehrnsberger R. 1998b — Eine neue Untergattung Haloseius subgen. nov. in der Gattung Halolaelaps Berlese & Trouessart, 1889 (Acari, Gamasida: Halolaelapidae) — Genus, 9: 421-429.
Blaszak C., Ehrnsberger R., Gwiazdowicz D.J. 2001 — Beschreibung des Männchens von Halolaelaps (Haloseius) sexclavatus (Oudemans, 1902) (Acari, Gamasida: Halolaelapidae) — Osnab. Nat. Mit., 27: 99-101.
Bregetova N.G., Shcherbak G.I. 1977 — Fam. Rhodacaridae - In: Gilyarov M.S., Bregetova N.G. (Eds) A Key to the Soil-Inhabiting Mites of the Mesostigmata — Nauka, Leningrad, p. 256-307. [In Russian]
Coulson S.J., Convey P., Aakra K., Aarvik L., Ávila-Jiménez M.L., Babenko A., Biersma E., Boström S., Brittain J., Carlsson A., Christoffersen K.S., De Smet W.H., Ekrem T., Fjellberg A., Füreder L., Gustafsson D., Gwiazdowicz D.J., Hansen L.O., Holmstrup M., Hullé M., Kaczmarek ., Kolicka M., Kuklin V., Lakka H-K., Lebedeva N., Makarova O., Maraldo K., Melekhina E., degaard F., Pilskog H.E., Simon J.C., Sohlenius B., Solh y T., S li G., Stur E., Tanasevitch A., Taskaeva A., Velle G., Zawierucha K., Zmudczy nska K. 2014 — The terrestrial and fresh water invertebrate biodiversity of the archipelagoes of the Barents Sea; Svalbard, Franz Josef Land and Novaya Zemlya — Soil Biol. Bioch., 68: 440-470.
Coulson S.J., Fjellberg A., Melekhina E.N., Taskaeva A.A., Lebedeva N.V., Belkina O., Seniczak S., Seniczak A., Gwiazdowicz D.J. 2015 — Microarthropod communities of industrially disturbed or imported soils in the High Arctic; the abandoned coal mining town of Pyramiden, Svalbard — Biodiversity and Conservation, 24: 1671-1690. doi:10.1007/s10531-015-0885-9 ![]()
Coulson S.J., Hodkinson I.D., Strathdee A.T., Block W., Webb N.R., Bale J.S., Worland M.R. 1995 — Thermal environments of Arctic soil organisms during winter — Arct. Alp. Res., 27: 365-371. doi:10.2307/1552029 ![]()
Evans G.O. 1963 — Observations on the chaetotaxy of the legs in the free-living Gamasina (Acari: Mesostigmata) — Bull. Brit. Mus. (Nat. Hist.), Zoology series, 10: 275-303.
Evans G.O., Till W.M. 1979 — Mesostigmatic mites of Britain and Ireland (Chelicerata: Acari-Parasitiformes). An introduction to their external morphology and classification — Trans. Zool. Soc. Lond., 35: 139-270. doi:10.1111/j.1096-3642.1979.tb00059.x ![]()
Gwiazdowicz, D.J. 2001 — Saprosecans bialoviensis sp. n. (Acari, Halolaelapidae) from Poland — Biologia, 56: 149-153.
Gwiazdowicz D.J., Gulvik M.E. 2008 — Mesostigmatid mites (Acari, Mesostigmata) in Svalbard — Proceedings of the 32nd International Polar Symposium, Wroclaw: 32-34.
Gwiazdowicz D.J., Teodorowicz E., Coulson S.J. 2011 — Redescription of Arctoseius haarlovi Lindquist, 1963 (Acari: Ascidae) from Spitsbergen — Entomol. Fen., 22: 140-148.
Halliday R.B. 2008 — Two new species of littoral mites from Australia (Acari: Halolaelapidae) — Syst. Appl. Acarol., 13: 214-230. doi:10.11158/saa.13.3.8 ![]()
Hirschmann W. 1966 — Die Gattung Halolaelaps Berlese & Trouessart 1889 nov. comb — Acarologie, 9: 21-24.
Hirschmann W., Götz H. 1968 — Neue Halolaelaps-Arten — Acarologie, 11: 7-10.
Karg W. 1993 — Acari (Acarina), Milben Parasitiformes (Anactinochaeta), Cohors Gamasina Leach. Raubmilben — Die Tierwelt Deutschlands, 59 Teil. Gustav Fischer Verlag, Jena, pp. 523.
Kolodochka L.A., Gwiazdowicz D.J. 2014 — A new species of predaceous mite of the genus Neoseiulus Hughes (Parasitiformes, Phytoseiidae), with redescriptions of N. magnanalis (Thor) and N. ellesmerei (Chant et Hansell), from Svalbard, High Arctic — Zootaxa, 3793: 441-452.
Lindquist E.E. 1994 — Some observations on the chaetotaxy of the caudal body region of Gamasine mites (Acari: Mesostigmata), with a modified notation for some ventrolateral body setae — Acarologia, 35: 323-326.
Lindquist E.E., Evans G.O. 1965 — Taxonomic concepts in the Ascidae, with a modified setal nomenclature for the idiosoma of the Gamasina (Acarina: Mesostigmata) — Mem. Entomol. Soc. Can., 47: 1-64. doi:10.4039/entm9747fv ![]()
Lindquist E.E., Krantz G.W., Walter D.E. 2009 — Order Mesostigmata — In: Krantz G.W., Walter D.E. (Eds) A Manual of Acarology, Third Edition. Texas Tech University Press, Lubbock, p. 124-232.
Norwegian Meteorological Institute 2012. ( http://retro.met.no/observasjoner/svalbard/SvalbardLufthavn/index.html ![]() . Accessed February 2012.)
. Accessed February 2012.)
This paper suffers from obviously erroneous setal notation on the female podonotal and opisthonotal shields, as well as not accounting for various of the shields’ pore-like structures.
Podonotal shield: setae denoted s2 through s6 are actually s1 through s5; the true s6 is a seta indicated as r6 on the corner of the shield, with true r6 on soft cuticle beside it.
Opisthonotal shield: seta on soft cuticle denoted Z5 is actually S5; the true Z5 is broken off (indicated only by its alveolus) on the right side, but is present on left side.
At least, illustrations of the legs are given, but it is unclear from what aspects, especially without any setal notation being given for perspective.
Further, under ‘Morphological variability”, comments are made regarding “Arctoseius haarlovi Lindquist 1963”, which is not cited in the references. In any case, that name is a nomen nudum, and the authors do not seem aware that the species was not validly described under that name until being done so as Arctoseius haarlovi Lindquist & Makarova, 2011, again not cited [Zool. Zhurnal 90: 923-941].



2016-08-24
Date accepted:
2016-10-26
Date published:
2017-04-20
Edited by:
Faraji, Farid

This work is licensed under a Creative Commons Attribution 4.0 International License
2017 Gwiazdowicz, Dariusz J. and Teodorowicz, Ewa
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)